

Germ-line mutations in p27Kip1 cause a multiple endocrine neoplasia syndrome in rats and humans
- PMID: 17030811
- PMCID: PMC1622862
- DOI: 10.1073/pnas.0603877103
Germ-line mutations in p27Kip1 cause a multiple endocrine neoplasia syndrome in rats and humans
Erratum in
- Proc Natl Acad Sci U S A. 2006 Dec 12;103(50):19213
Abstract
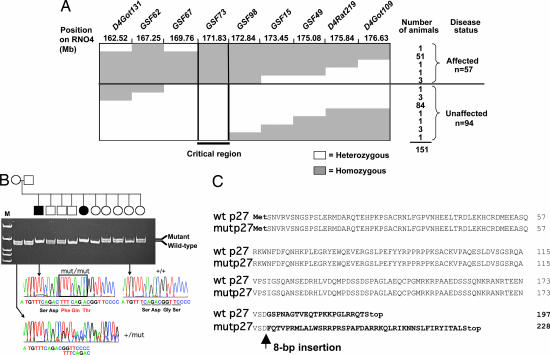
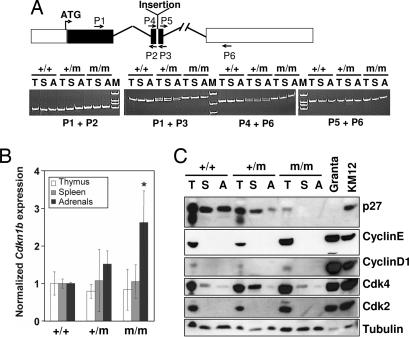
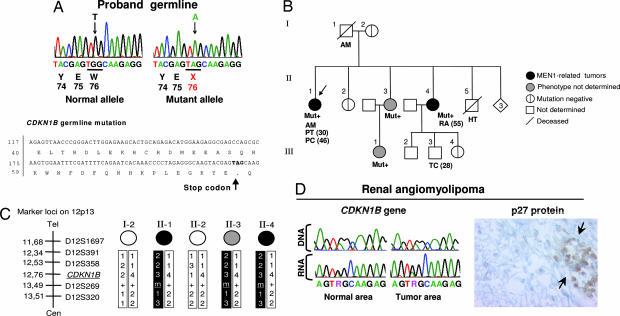
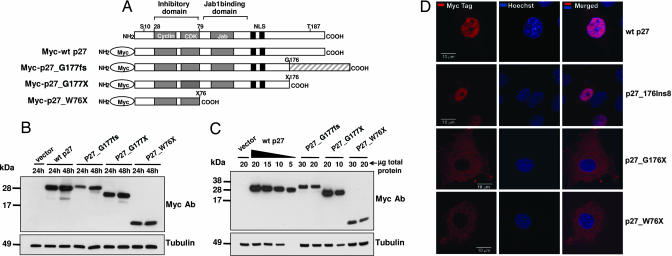
MENX is a recessive multiple endocrine neoplasia-like syndrome in the rat. The tumor spectrum in MENX overlaps those of human multiple endocrine neoplasia (MEN) types 1 and 2. We mapped the MenX locus to the distal part of rat chromosome 4, excluding the homologs of the genes responsible for the MEN syndromes (RET and MEN1) and syndromes with an endocrine tumor component (VHL and NF1). We report the fine mapping of the disease locus and the identification of a homozygous frameshift mutation in Cdkn1b, encoding the cyclin-dependent kinase inhibitor p27(Kip1). As a consequence of the mutation, MENX-affected rats show dramatic reduction in p27(Kip1) protein. We have identified a germ-line nonsense mutation in the human CDKN1B gene in a MEN1 mutation-negative patient presenting with pituitary and parathyroid tumors. Expanded pedigree analysis shows that the mutation is associated with the development of an MEN1-like phenotype in multiple generations. Our findings demonstrate that germ-line mutations in p27(Kip1) can predispose to the development of multiple endocrine tumors in both rats and humans.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Fritz A, Walch A, Piotrowska K, Rosemann M, Schaffer E, Weber K, Timper A, Wildner G, Graw J, Hoefler H, et al. Cancer Res. 2002;62:3048–3051. - PubMed
-
- Piotrowska K, Pellegata NS, Rosemann M, Fritz A, Graw J, Atkinson MJ. Mamm Genome. 2004;15:135–141. - PubMed
-
- Namihira H, Sato M, Matsubara S, Ohye H, Bhuiyan M, Murao K, Takahara J. Endocr J. 1999;46:811–816. - PubMed
-
- Sakurai A, Katai M, Yumita W, Minemura K, Hashizume K. Endocrine. 2004;23:45–49. - PubMed
-
- Nakayama K, Ishida N, Shirane M, Inomata A, Inoue T, Shishido N, Horii I, Loh DY, Nakayama K. Cell. 1996;85:707–720. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
