

Myostatin modulates adipogenesis to generate adipocytes with favorable metabolic effects
- PMID: 17030820
- PMCID: PMC1592529
- DOI: 10.1073/pnas.0607501103
Myostatin modulates adipogenesis to generate adipocytes with favorable metabolic effects
Abstract
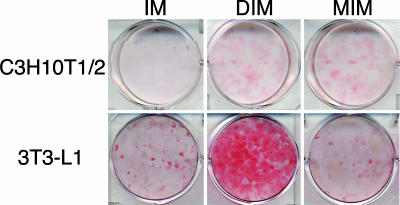
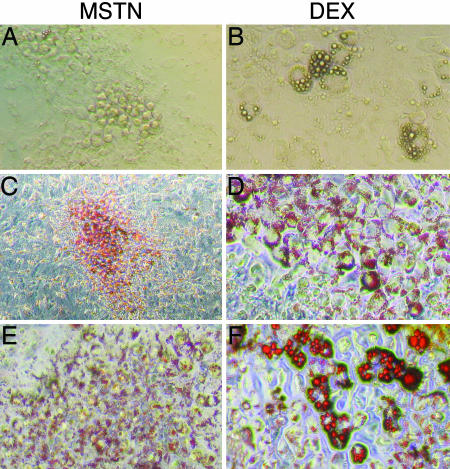
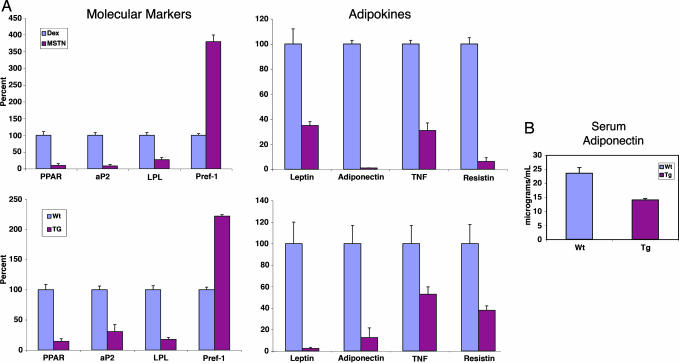
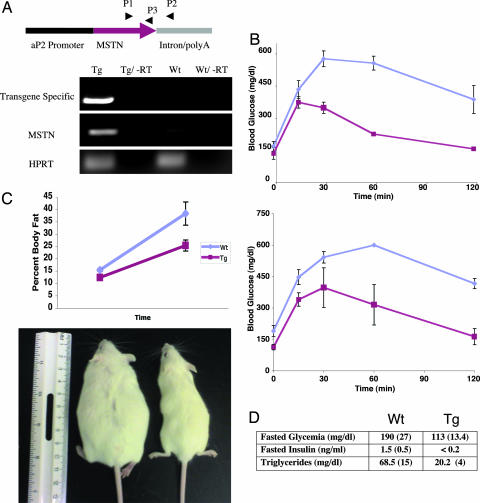
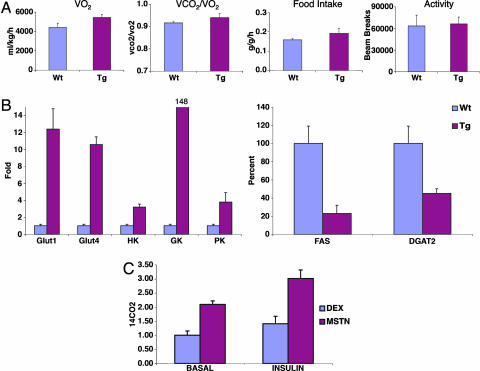
A pluripotent cell line, C3H10T1/2, is induced to undergo adipogenesis by a mixture of factors that includes a glucocorticoid such as dexamethasone. We found that expression of myostatin (MSTN), a TGF-beta family member extensively studied in muscle, was induced by dexamethasone under those differentiation conditions. Moreover, MSTN could substitute for dexamethasone in the adipogenesis mixture. However, the adipocytes induced by MSTN in both cell culture and transgenic mice were small and expressed markers characteristic of immature adipocytes. These adipocytes exhibited cell-autonomous increases in insulin sensitivity and glucose oxidation. In mice, these effects produced elevated systemic insulin sensitivity and resistance to diet-induced obesity. Modulation of the final stages of adipogenesis may provide a novel approach to understanding and treating metabolic disease.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Shepherd PR, Gnudi L, Tozzo E, Yang H, Leach F, Kahn BB. J Biol Chem. 1993;268:22243–22246. - PubMed
-
- Kershaw EE, Flier JS. J Clin Endocrinol Metab. 2004;89:2548–2556. - PubMed
-
- Yu ZK, Wright JT, Hausman GJ. Obes Res. 1997;5:9–15. - PubMed
-
- MacDougald OA, Mandrup S. Trends Endocrinol Metab. 2002;13:5–11. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
