

Mapping the assembly pathways that specify formation of the trilaminar kinetochore plates in human cells
- PMID: 17030981
- PMCID: PMC2064494
- DOI: 10.1083/jcb.200606020
Mapping the assembly pathways that specify formation of the trilaminar kinetochore plates in human cells
Abstract
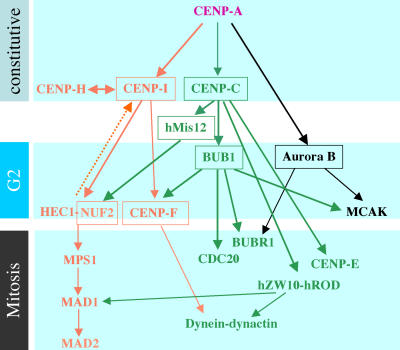
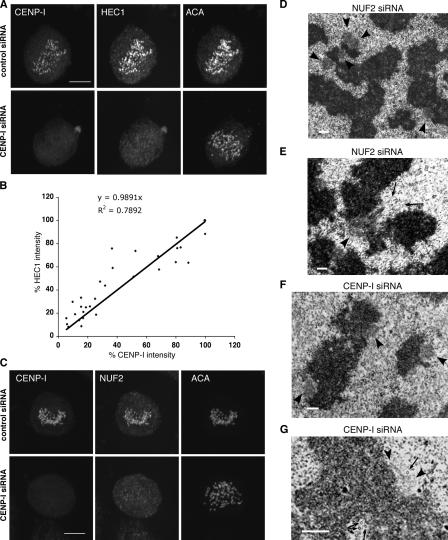
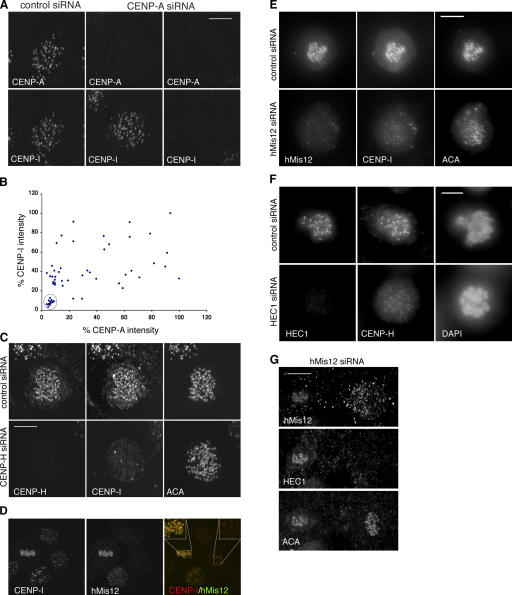
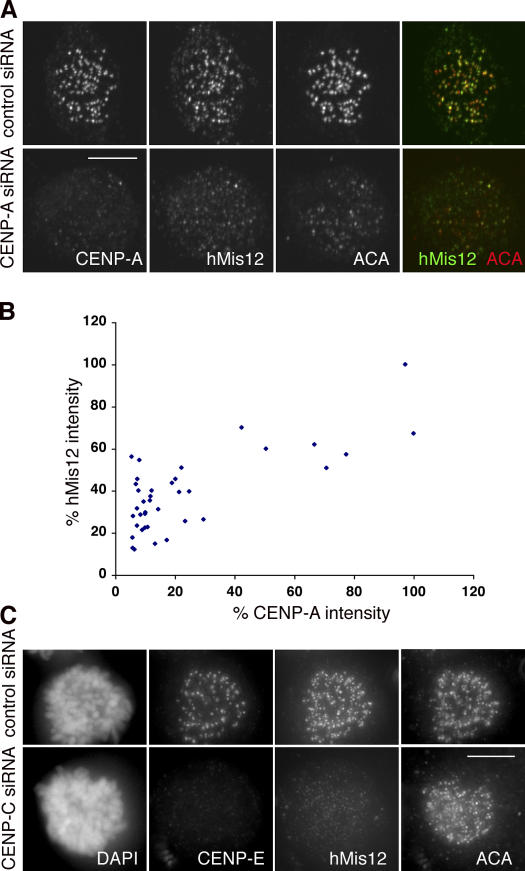
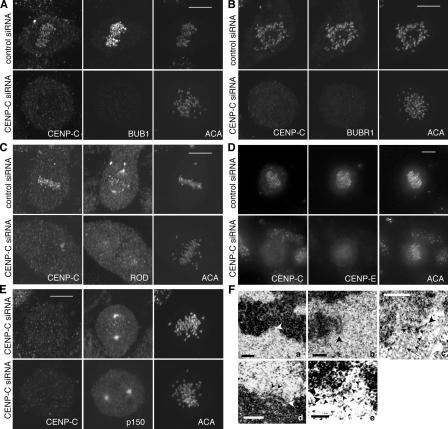
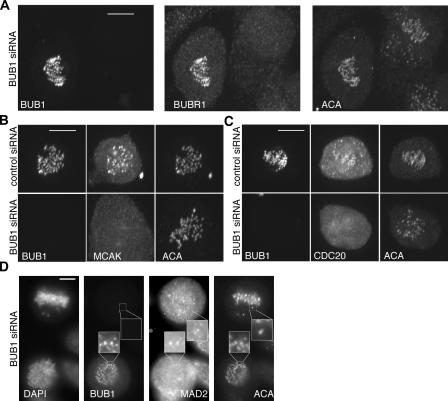
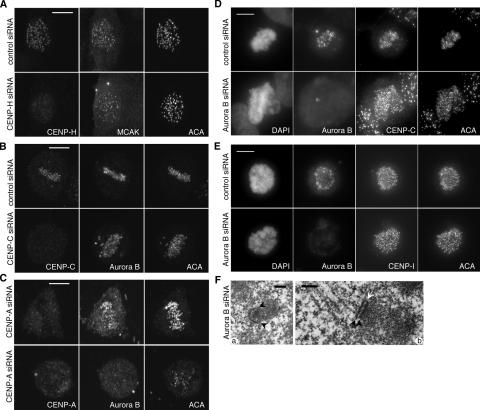
We report the interactions amongst 20 proteins that specify their assembly to the centromere-kinetochore complex in human cells. Centromere protein (CENP)-A is at the top of a hierarchy that directs three major pathways, which are specified by CENP-C, -I, and Aurora B. Each pathway consists of branches that intersect to form nodes that may coordinate the assembly process. Complementary EM studies found that the formation of kinetochore trilaminar plates depends on the CENP-I/NUF2 branch, whereas CENP-C and Aurora B affect the size, shape, and structural integrity of the plates. We found that hMis12 is not constitutively localized at kinetochores, and that it is not essential for recruiting CENP-I. Our studies also revealed that kinetochores in HeLa cells contain an excess of CENP-A, of which approximately 10% is sufficient to promote the assembly of normal levels of kinetochore proteins. We elaborate on a previous model that suggested kinetochores are assembled from repetitive modules (Zinkowski, R.P., J. Meyne, and B.R. Brinkley. 1991. J. Cell Biol. 113:1091-110).
Figures
References
-
- Adams, R.R., M. Carmena, and W.C. Earnshaw. 2001. Chromosomal passengers and the (aurora) ABCs of mitosis. Trends Cell Biol. 11:49–54. - PubMed
-
- Amor, D.J., P. Kalitsis, H. Sumer, and K.H. Choo. 2004. Building the centromere: from foundation proteins to 3D organization. Trends Cell Biol. 14:359–368. - PubMed
-
- Andrews, P.D., Y. Ovechkina, N. Morrice, M. Wagenbach, K. Duncan, L. Wordeman, and J.R. Swedlow. 2004. Aurora B regulates MCAK at the mitotic centromere. Dev. Cell. 6:253–268. - PubMed
-
- Bharadwaj, R., W. Qi, and H. Yu. 2004. Identification of two novel components of the human NDC80 kinetochore complex. J. Biol. Chem. 279:13076–13085. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
