

Interplay between PIP3 and calmodulin regulation of olfactory cyclic nucleotide-gated channels
- PMID: 17032767
- PMCID: PMC1622874
- DOI: 10.1073/pnas.0603344103
Interplay between PIP3 and calmodulin regulation of olfactory cyclic nucleotide-gated channels
Abstract
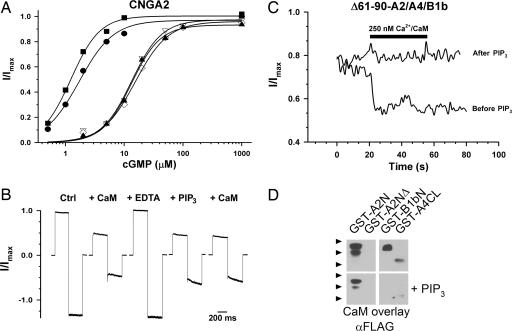
Phosphatidylinositol-3,4,5-trisphosphate (PIP3) has been proposed to modulate the odorant sensitivity of olfactory sensory neurons by inhibiting activation of cyclic nucleotide-gated (CNG) channels in the cilia. When applied to the intracellular face of excised patches, PIP3 has been shown to inhibit activation of heteromeric olfactory CNG channels, composed of CNGA2, CNGA4, and CNGB1b subunits, and homomeric CNGA2 channels. In contrast, we discovered that channels formed by CNGA3 subunits from cone photoreceptors were unaffected by PIP3. Using chimeric channels and a deletion mutant, we determined that residues 61-90 within the N terminus of CNGA2 are necessary for PIP3 regulation, and a biochemical "pulldown" assay suggests that PIP3 directly binds this region. The N terminus of CNGA2 contains a previously identified calcium-calmodulin (Ca2+/CaM)-binding domain (residues 68-81) that mediates Ca2+/CaM inhibition of homomeric CNGA2 channels but is functionally silent in heteromeric channels. We discovered, however, that this region is required for PIP3 regulation of both homomeric and heteromeric channels. Furthermore, PIP3 occluded the action of Ca2+/CaM on both homomeric and heteromeric channels, in part by blocking Ca2+/CaM binding. Our results establish the importance of the CNGA2 N terminus for PIP3 inhibition of olfactory CNG channels and suggest that PIP3 inhibits channel activation by disrupting an autoexcitatory interaction between the N and C termini of adjacent subunits. By dramatically suppressing channel currents, PIP3 may generate a shift in odorant sensitivity that does not require prior channel activity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
