

Inhibitory effect of gamma interferon on BK virus gene expression and replication
- PMID: 17035315
- PMCID: PMC1797268
- DOI: 10.1128/JVI.01571-06
Inhibitory effect of gamma interferon on BK virus gene expression and replication
Abstract
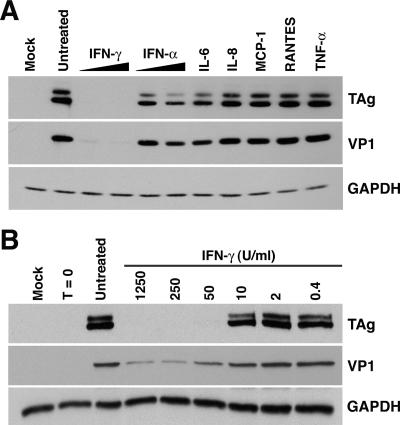
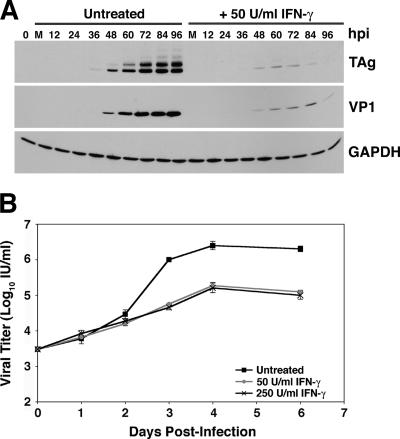
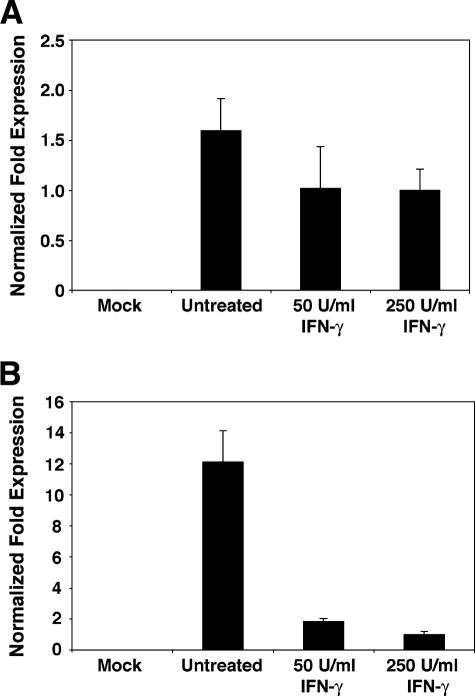
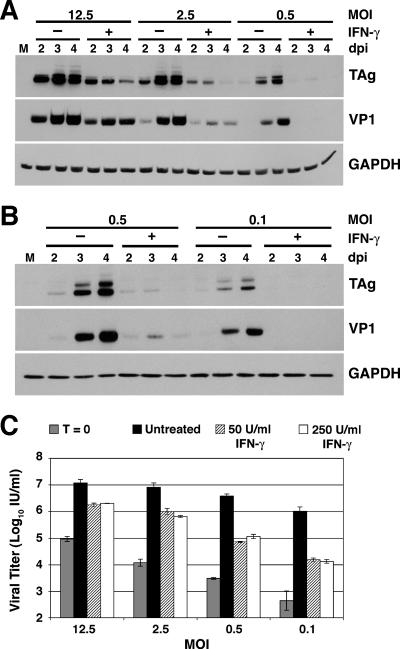
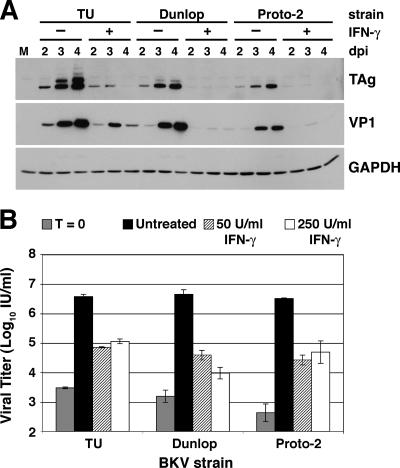
BK virus (BKV) is widely accepted to be the causative agent of polyomavirus nephropathy. In immunocompromised individuals, especially kidney transplant recipients, BKV can replicate in kidney epithelial cells, causing loss of renal function and eventual destruction of the graft. Advances in immunosuppressive therapies may be partially responsible for the increasing incidence of polyomavirus nephropathy among transplant recipients by more effectively eliminating components of the immune system, such as gamma interferon (IFN-gamma)-producing lymphocytes, that keep BKV infections at a subclinical level. In this study, we investigated the role of IFN-gamma in regulating lytic infection by BKV. Treatment with IFN-gamma inhibited the expression of the viral early protein large tumor antigen (TAg) and the late protein VP1 in a dose-dependent manner. We detected 1.6- and 12-fold reductions in TAg transcripts at 48 and 96 h postinfection, respectively, with 250 U/ml IFN-gamma, suggesting that IFN-gamma-mediated inhibition occurs at the level of transcription. Furthermore, IFN-gamma inhibited the level of viral progeny production as much as 50-fold at a multiplicity of infection (MOI) of 0.5 and 80-fold at an MOI of 0.1. The inhibitory effects of IFN-gamma were similar for three different strains of BKV examined. These results indicate an important role for IFN-gamma in regulating BKV lytic infection.
Figures
References
-
- Bohl, D. L., G. A. Storch, C. Ryschkewitsch, M. Gaudreault-Keener, M. A. Schnitzler, E. O. Major, and D. C. Brennan. 2005. Donor origin of BK virus in renal transplantation and role of HLA C7 in susceptibility to sustained BK viremia. Am. J. Transplant. 5:2213-2221. - PubMed
-
- Briggs, J. P., W. Kriz, and J. B. Schnermann. 2001. Overview of renal function structure, p. 3-19. In A. Greenberg et al. (ed.), Primer on kidney diseases, 3rd ed. Academic Press, New York, NY.
-
- Chen, Y., J. Trofe, J. Gordon, R. A. Du Pasquier, P. Roy-Chaudhury, M. J. Kuroda, E. S. Woodle, K. Khalili, and I. J. Koralnik. 2006. Interplay of cellular and humoral immune responses against BK virus in kidney transplant recipients with polyomavirus nephropathy. J. Virol. 80:3495-3505. - PMC - PubMed
-
- Chesters, P. M., J. Heritage, and D. J. McCance. 1983. Persistence of DNA sequences of BK virus and JC virus in normal human tissues and in diseased tissues. J. Infect. Dis. 147:676-684. - PubMed
-
- Comoli, P., A. Azzi, R. Maccario, S. Basso, G. Botti, G. Basile, I. Fontana, M. Labirio, A. Cometa, F. Poli, F. Perfumo, F. Locatelli, and F. Ginevri. 2004. Polyomavirus BK-specific immunity after kidney transplantation. Transplantation 78:1229-1232. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
