

Evolution of the Rho family of ras-like GTPases in eukaryotes
- PMID: 17035353
- PMCID: PMC2665304
- DOI: 10.1093/molbev/msl145
Evolution of the Rho family of ras-like GTPases in eukaryotes
Abstract
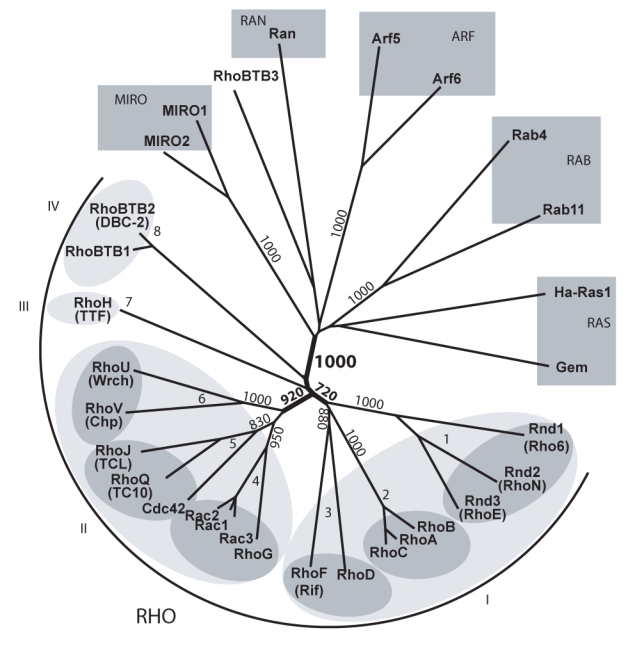
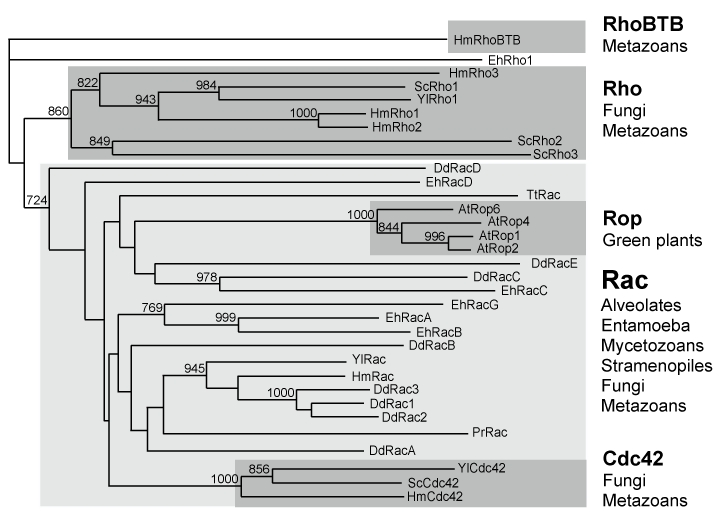
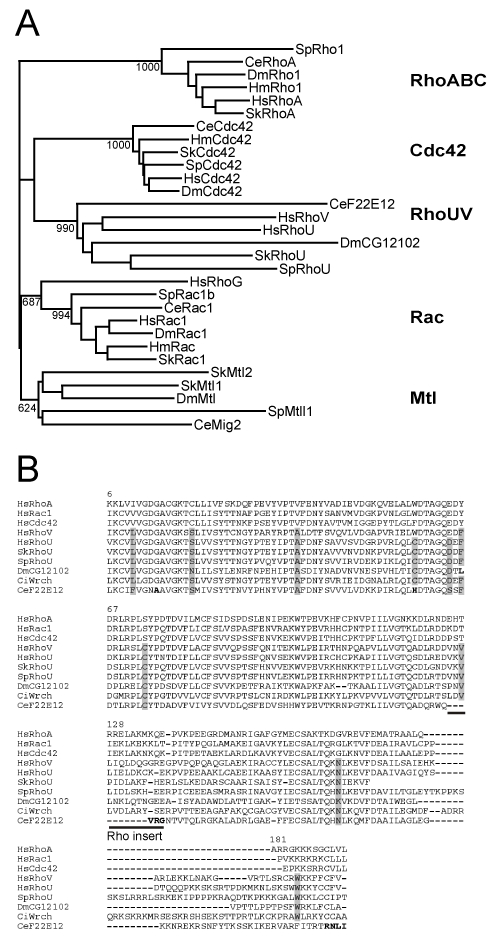
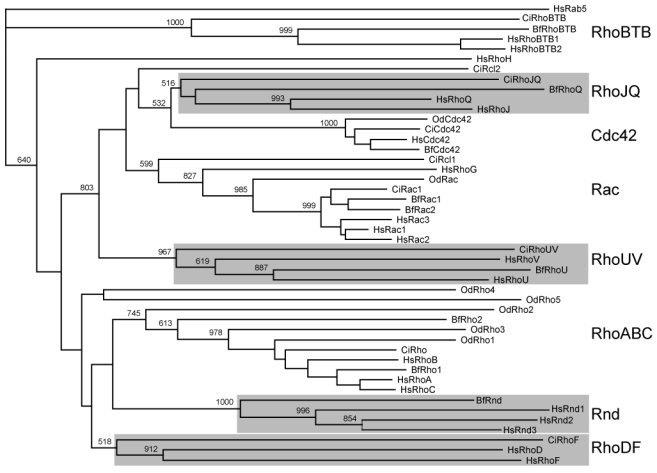
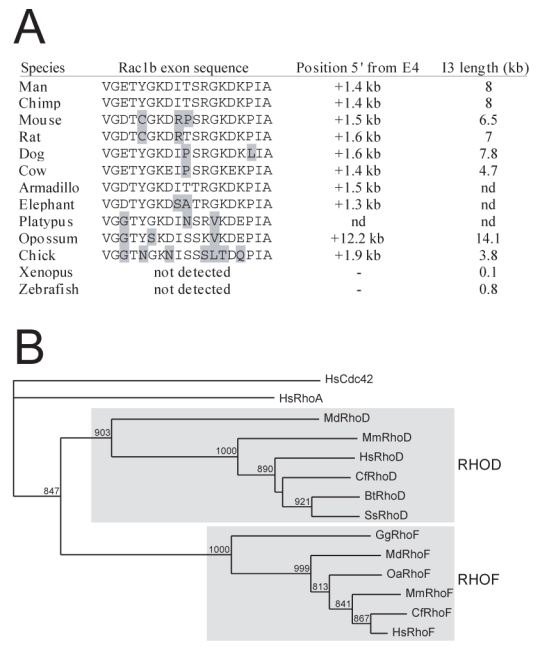
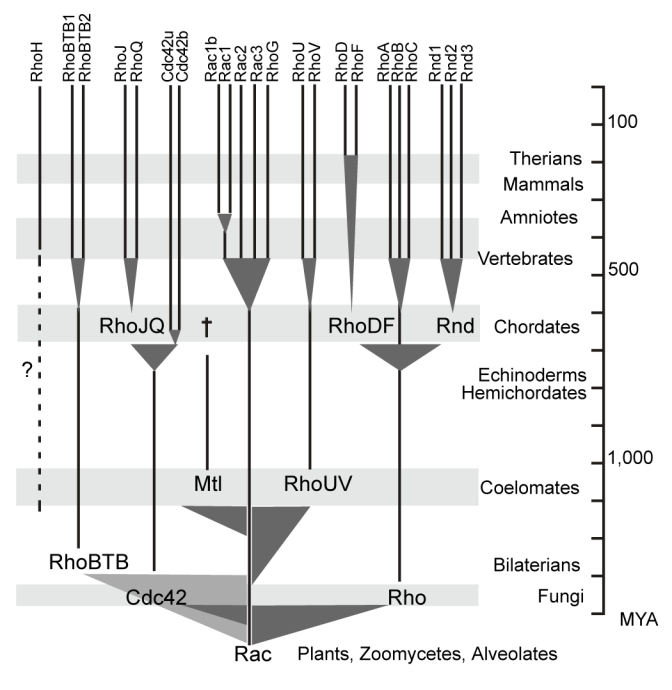
GTPases of the Rho family are molecular switches that play important roles in converting and amplifying external signals into cellular effects. Originally demonstrated to control the dynamics of the F-actin cytoskeleton, Rho GTPases have been implicated in many basic cellular processes that influence cell proliferation, differentiation, motility, adhesion, survival, or secretion. To elucidate the evolutionary history of the Rho family, we have analyzed over 20 species covering major eukaryotic clades from unicellular organisms to mammals, including platypus and opossum, and have reconstructed the ontogeny and the chronology of emergence of the different subfamilies. Our data establish that the 20 mammalian Rho members are structured into 8 subfamilies, among which Rac is the founder of the whole family. Rho, Cdc42, RhoUV, and RhoBTB subfamilies appeared before Coelomates and RhoJQ, Cdc42 isoforms, RhoDF, and Rnd emerged in chordates. In vertebrates, gene duplications and retrotranspositions increased the size of each chordate Rho subfamily, whereas RhoH, the last subfamily, arose probably by horizontal gene transfer. Rac1b, a Rac1 isoform generated by alternative splicing, emerged in amniotes, and RhoD, only in therians. Analysis of Rho mRNA expression patterns in mouse tissues shows that recent subfamilies have tissue-specific and low-level expression that supports their implication only in narrow time windows or in differentiated metabolic functions. These findings give a comprehensive view of the evolutionary canvas of the Rho family and provide guides for future structure and evolution studies of other components of Rho signaling pathways, in particular regulators of the RhoGEF family.
Figures
References
-
- Abe T, Kato M, Miki H, Takenawa T, Endo T. Small GTPase Tc10 and its homologue RhoT induce N-WASP-mediated long process formation and neurite outgrowth. Journal of Cell Science. 2003;116:155–168. - PubMed
-
- Aktories K, Barbieri JT. Bacterial cytotoxins: targeting eukaryotic switches. Nat Rev Microbiol. 2005;3:397–410. - PubMed
-
- Andersen PR, Devare SG, Tronick SR, Ellis RW, Aaronson SA, Scolnick EM. Generation of BALB-MuSV and Ha-MuSC by type C virus transduction of homologous transforming genes from different species. Cell. 1981;26:129–134. - PubMed
-
- Aronheim A, Broder YC, Cohen A, Fritsch A, Belisle B, Abo A. Chp, a homologue of the GTPase Cdc42Hs, activates the JNK pathway and is implicated in reorganizing the actin cytoskeleton. Current Biology. 1998;8:1125–1128. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
