

Epigenetic determination of a cell-specific gene expression program by ATF-2 and the histone variant macroH2A
- PMID: 17036053
- PMCID: PMC1618095
- DOI: 10.1038/sj.emboj.7601364
Epigenetic determination of a cell-specific gene expression program by ATF-2 and the histone variant macroH2A
Abstract
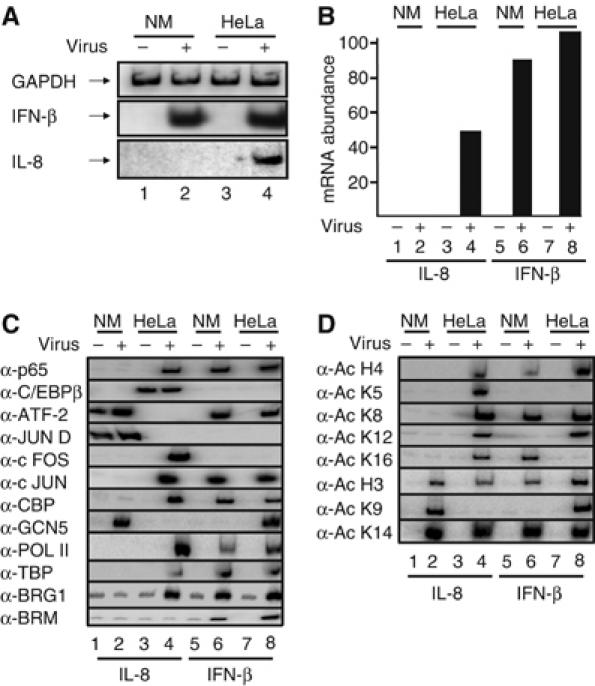
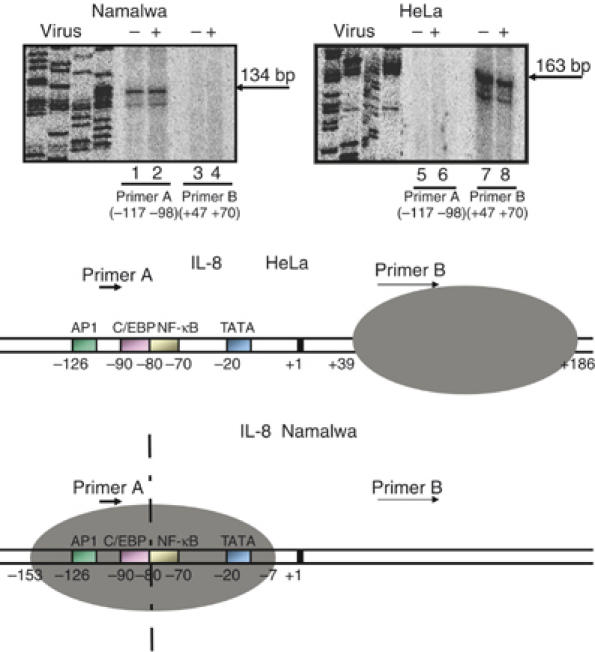
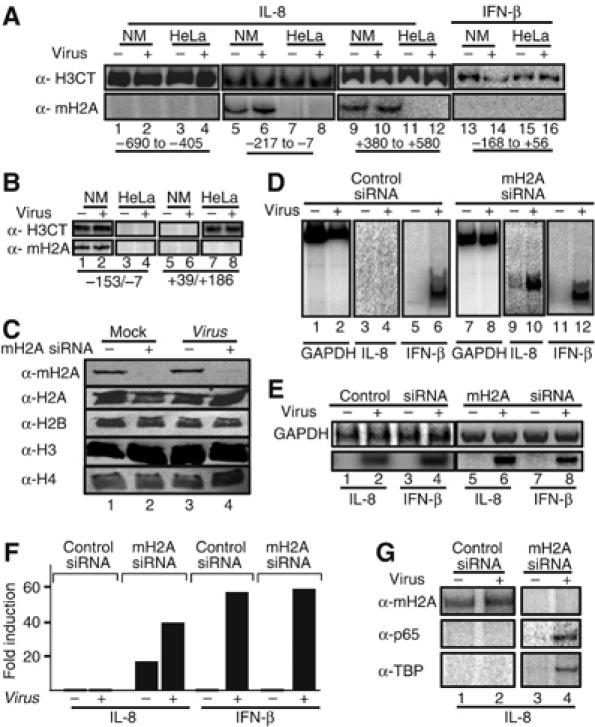
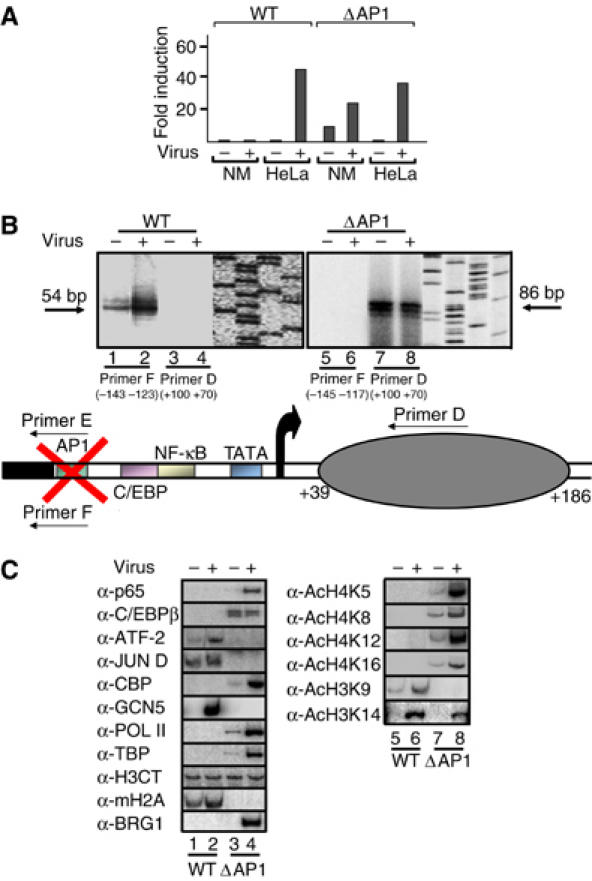
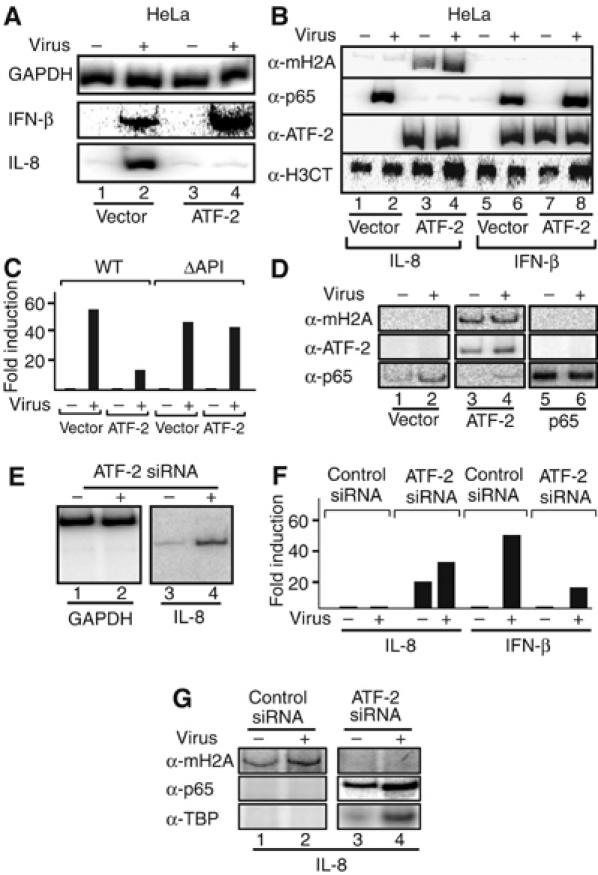
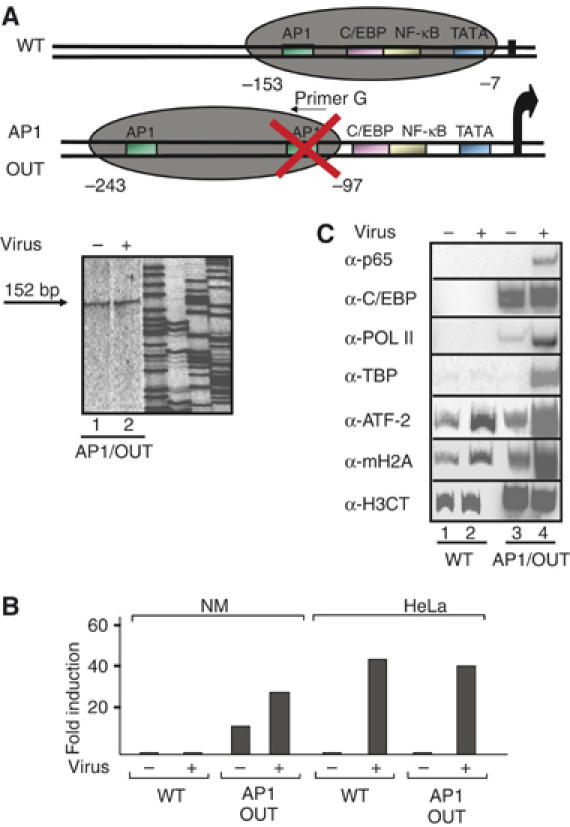
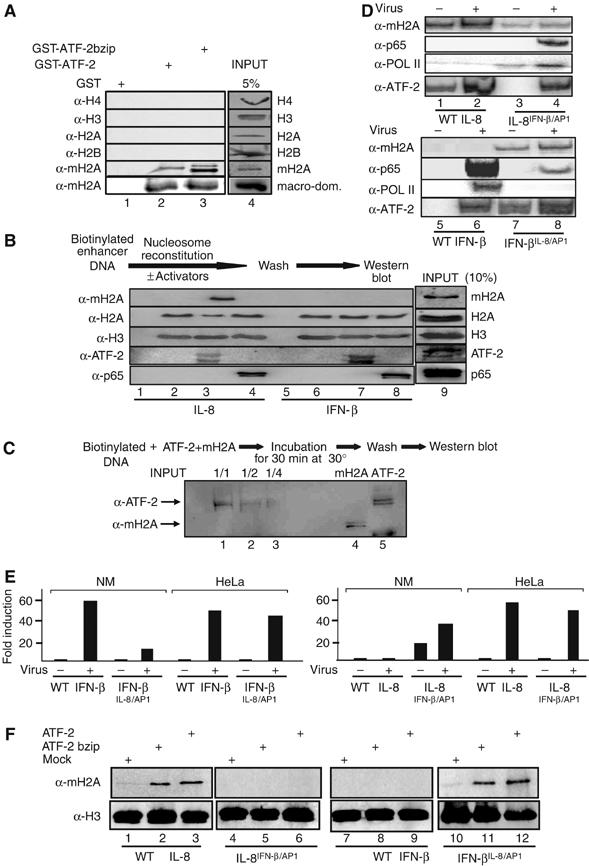
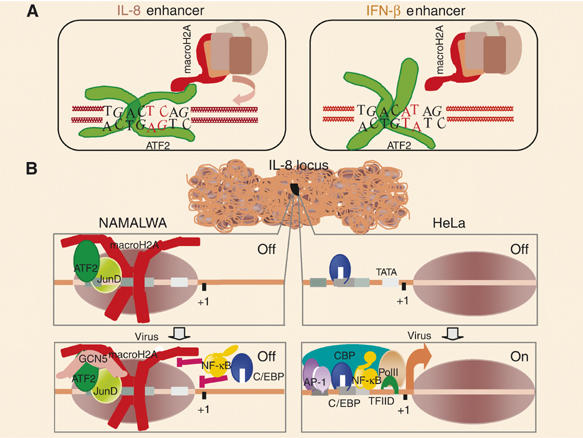
Transcriptional activation of the interleukin-8 (IL-8) gene is restricted to distinct cell types, although the transcriptional regulatory proteins controlling IL-8 gene expression are ubiquitous. We show that cell-specific transcription of IL-8 is due to the distinct chromatin architecture on the enhancer/promoter before the arrival of the inducing signal. In expressing epithelial cells the enhancer/promoter is nucleosome-free, whereas in non-expressing B cells a nucleosome masks the entire regulatory region. The B-cell-specific nucleosome contains the histone variant macroH2A, which is responsible for preventing transcription factor binding. Recruitment of the repressive macroH2A nucleosome requires direct interactions between ATF-2 bound to the nearby AP1 site and macroH2A and it is regulated by DNA-induced protein allostery. siRNA against ATF-2 or macroH2A rescues IL-8 transcription in B cells. Thus, a transcription factor can work as a transcriptional repressor by orchestrating and maintaining the assembly of specialized local chromatin architectures.
Figures
References
-
- Abbott DW, Chadwick BP, Thambirajah A, Ausio J (2005) Beyond the Xi: macroH2A chromatin distribution and post-translational modification in an avian system. J Biol Chem 280: 16437–16445 - PubMed
-
- Abbott DW, Laszczak M, Lewis JD, Su H, Moore SC, Hills M, Dimitrov S, Ausio J (2004) Structural characterization of macroH2A containing chromatin. Biochemistry 43: 1352–1359 - PubMed
-
- Agalioti T, Chen G, Thanos D (2002) Deciphering the transcriptional histone acetylation code for a human gene. Cell 111: 381–392 - PubMed
-
- Angelov D, Molla A, Perche PY, Hans F, Cote J, Khochbin S, Bouvet P, Dimitrov S (2003) The histone variant macroH2A interferes with transcription factor binding and SWI/SNF nucleosome remodeling. Mol Cell 11: 1033–1041 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
