

Independent centromere formation in a capricious, gene-free domain of chromosome 13q21 in Old World monkeys and pigs
- PMID: 17040560
- PMCID: PMC1794570
- DOI: 10.1186/gb-2006-7-10-r91
Independent centromere formation in a capricious, gene-free domain of chromosome 13q21 in Old World monkeys and pigs
Abstract
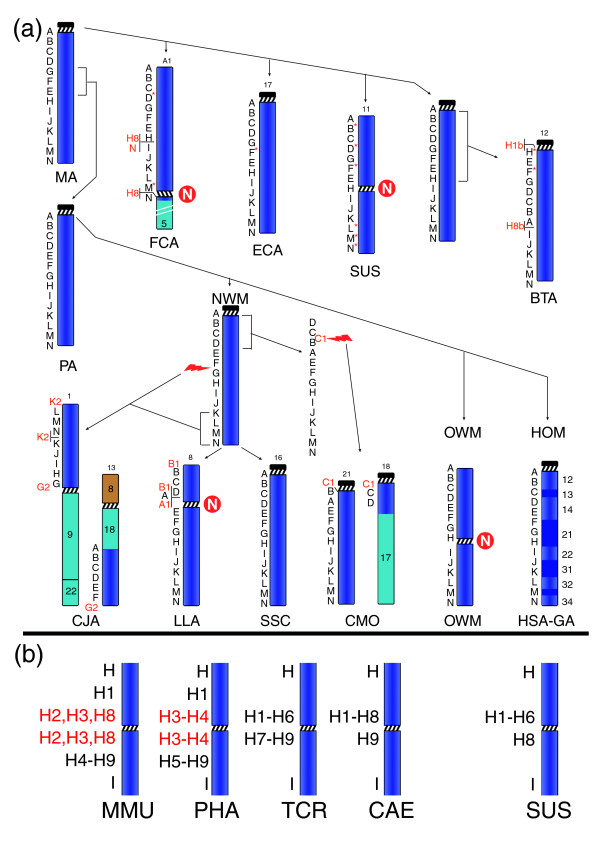
Background: Evolutionary centromere repositioning and human analphoid neocentromeres occurring in clinical cases are, very likely, two stages of the same phenomenon whose properties still remain substantially obscure. Chromosome 13 is the chromosome with the highest number of neocentromeres. We reconstructed the mammalian evolutionary history of this chromosome and characterized two human neocentromeres at 13q21, in search of information that could improve our understanding of the relationship between evolutionarily new centromeres, inactivated centromeres, and clinical neocentromeres.
Results: Chromosome 13 evolution was studied, using FISH experiments, across several diverse superordinal phylogenetic clades spanning >100 million years of evolution. The analysis revealed exceptional conservation among primates (hominoids, Old World monkeys, and New World monkeys), Carnivora (cat), Perissodactyla (horse), and Cetartiodactyla (pig). In contrast, the centromeres in both Old World monkeys and pig have apparently repositioned independently to a central location (13q21). We compared these results to the positions of two human 13q21 neocentromeres using chromatin immunoprecipitation and genomic microarrays.
Conclusion: We show that a gene-desert region at 13q21 of approximately 3.9 Mb in size possesses an inherent potential to form evolutionarily new centromeres over, at least, approximately 95 million years of mammalian evolution. The striking absence of genes may represent an important property, making the region tolerant to the extensive pericentromeric reshuffling during subsequent evolution. Comparison of the pericentromeric organization of chromosome 13 in four Old World monkey species revealed many differences in sequence organization. The region contains clusters of duplicons showing peculiar features.
Figures


References
-
- Warburton PE, Dolled M, Mahmood R, Alonso A, Li S, Naritomi K, Tohma T, Nagai T, Hasegawa T, Ohashi H, et al. Molecular cytogenetic analysis of eight inversion duplications of human chromosome 13q that each contain a neocentromere. Am J Hum Genet. 2000;66:1794–1806. doi: 10.1086/302924. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
