

Reduction of anion reversal potential subverts the inhibitory control of firing rate in spinal lamina I neurons: towards a biophysical basis for neuropathic pain
- PMID: 17040565
- PMCID: PMC1624821
- DOI: 10.1186/1744-8069-2-32
Reduction of anion reversal potential subverts the inhibitory control of firing rate in spinal lamina I neurons: towards a biophysical basis for neuropathic pain
Abstract
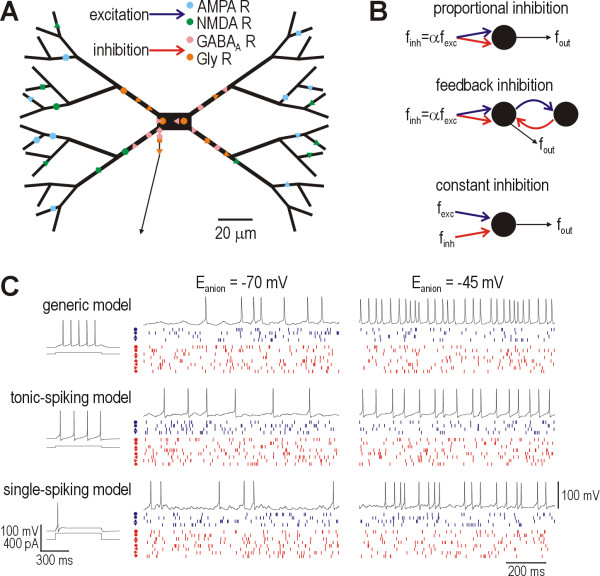
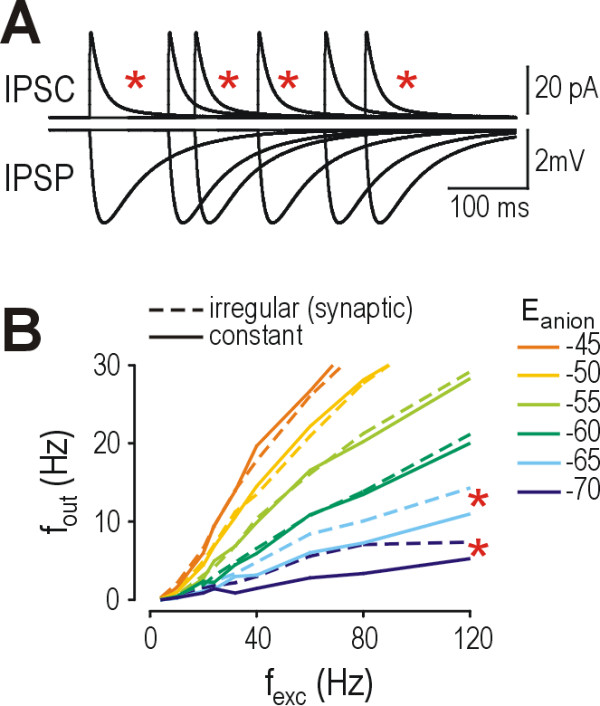
Background: Reduction of the transmembrane chloride gradient in spinal lamina I neurons contributes to the cellular hyperexcitability producing allodynia and hyperalgesia after peripheral nerve injury. The resultant decrease in anion reversal potential (i.e. shift in Eanion to less negative potentials) reduces glycine/GABAA receptor-mediated hyperpolarization, but the large increase in membrane conductance caused by inhibitory input can nonetheless shunt concurrent excitatory input. Without knowing the relative contribution of hyperpolarization and shunting to inhibition's modulation of firing rate, it is difficult to predict how much net disinhibition results from reduction of Eanion. We therefore used a biophysically accurate lamina I neuron model to investigate quantitatively how changes in Eanion affect firing rate modulation.
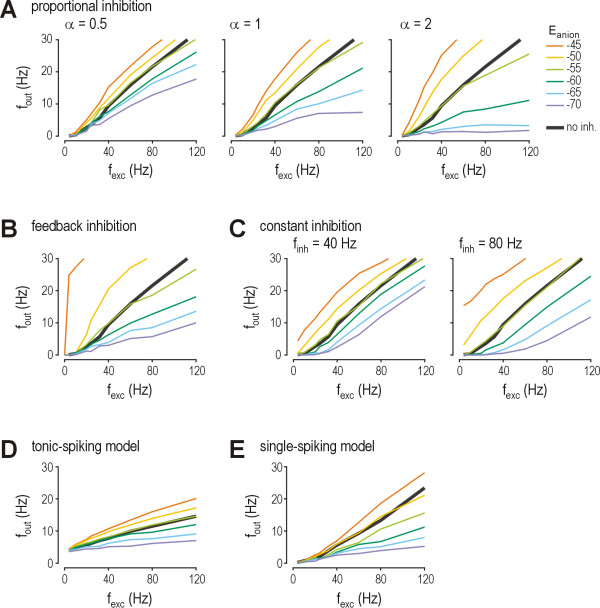
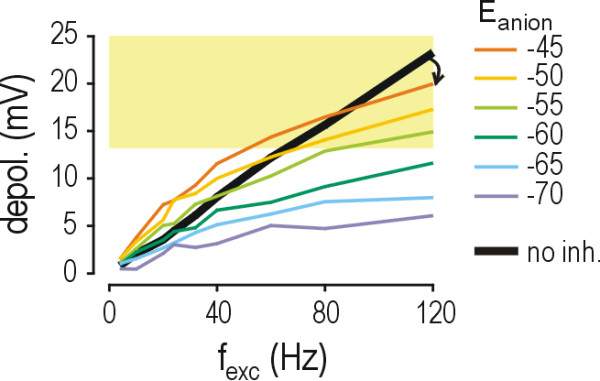
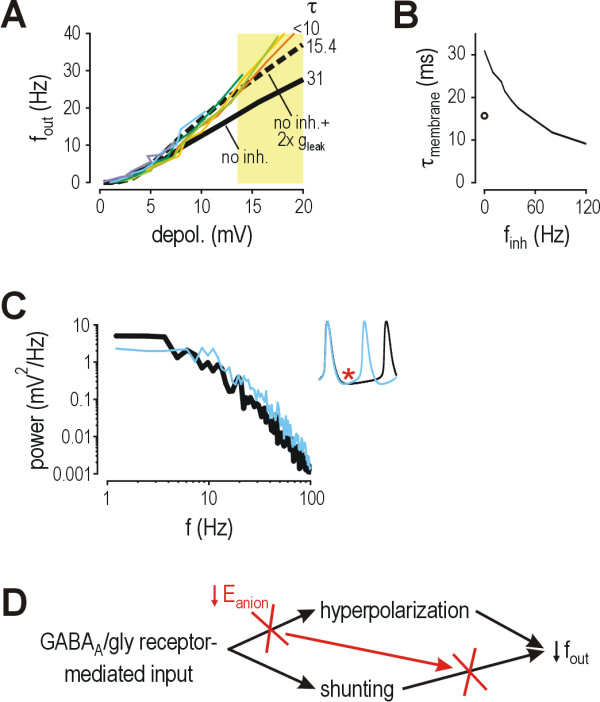
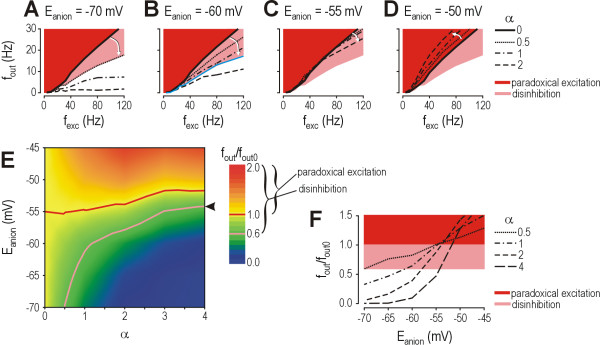
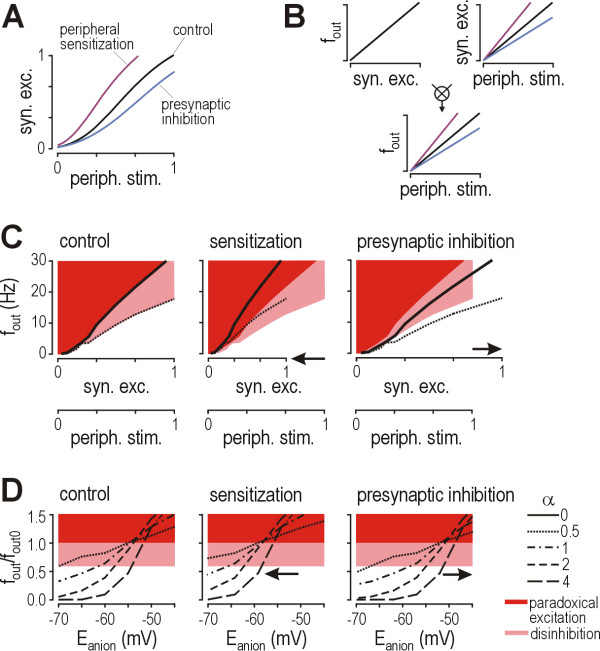
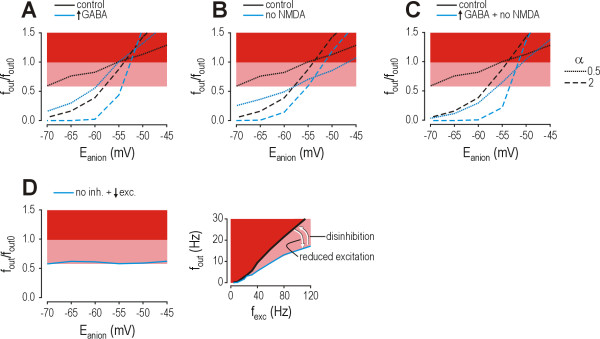
Results: Simulations reveal that even a small reduction of Eanion compromises inhibitory control of firing rate because reduction of Eanion not only decreases glycine/GABAA receptor-mediated hyperpolarization, but can also indirectly compromise the capacity of shunting to reduce spiking. The latter effect occurs because shunting-mediated modulation of firing rate depends on a competition between two biophysical phenomena: shunting reduces depolarization, which translates into reduced spiking, but shunting also shortens the membrane time constant, which translates into faster membrane charging and increased spiking; the latter effect predominates when average depolarization is suprathreshold. Disinhibition therefore occurs as both hyperpolarization- and shunting-mediated modulation of firing rate are subverted by reduction of Eanion. Small reductions may be compensated for by increased glycine/GABAA receptor-mediated input, but the system decompensates (i.e. compensation fails) as reduction of Eanion exceeds a critical value. Hyperexcitability necessarily develops once disinhibition becomes incompensable. Furthermore, compensation by increased glycine/GABAA receptor-mediated input introduces instability into the system, rendering it increasingly prone to abrupt decompensation and even paradoxical excitation.
Conclusion: Reduction of Eanion dramatically compromises the inhibitory control of firing rate and, if compensation fails, is likely to contribute to the allodynia and hyperalgesia associated with neuropathic pain. These data help explain the relative intractability of neuropathic pain and illustrate how it is important to choose therapies not only based on disease mechanism, but based on quantitative understanding of that mechanism.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
