

SIZ1 small ubiquitin-like modifier E3 ligase facilitates basal thermotolerance in Arabidopsis independent of salicylic acid
- PMID: 17041025
- PMCID: PMC1676064
- DOI: 10.1104/pp.106.088831
SIZ1 small ubiquitin-like modifier E3 ligase facilitates basal thermotolerance in Arabidopsis independent of salicylic acid
Abstract
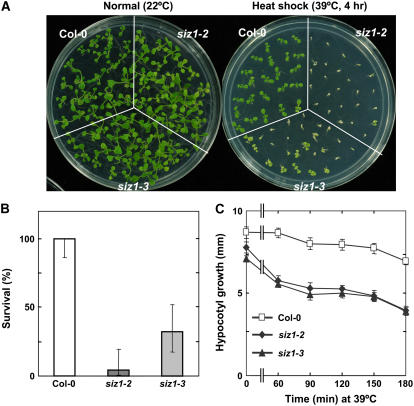
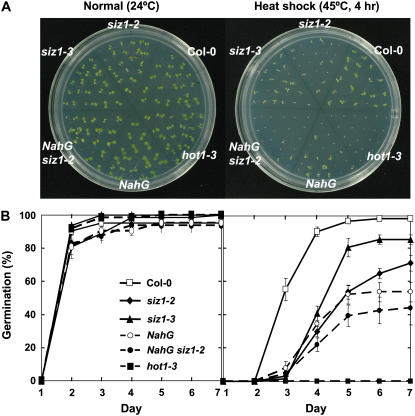
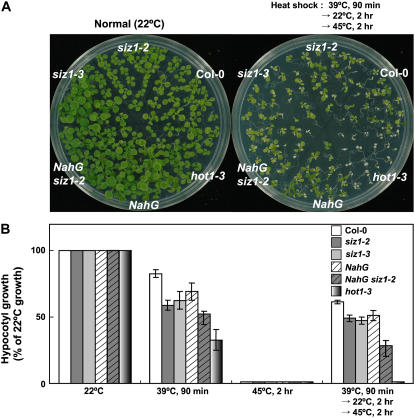
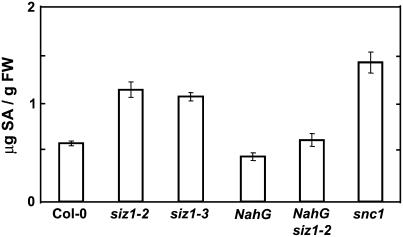
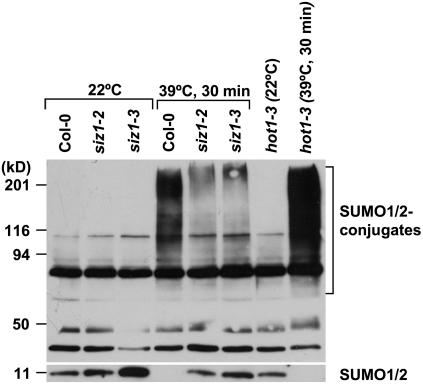
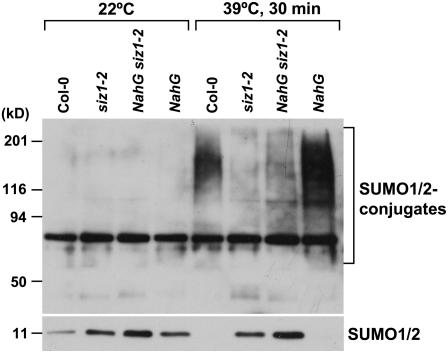
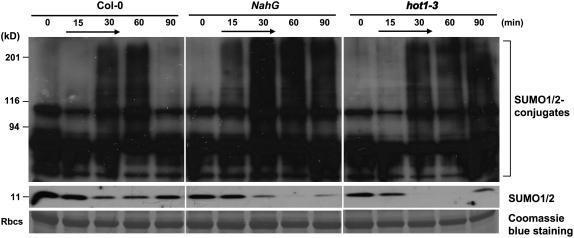
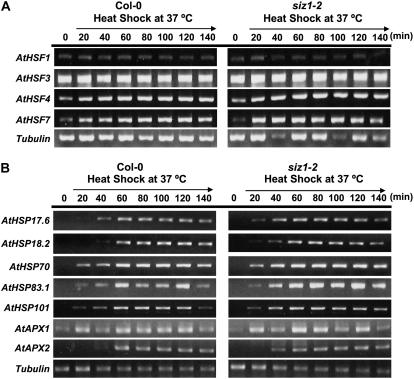
Small ubiquitin-like modifier (SUMO) conjugation/deconjugation to heat shock transcription factors regulates DNA binding of the peptides and activation of heat shock protein gene expression that modulates thermal adaptation in metazoans. SIZ1 is a SUMO E3 ligase that facilitates SUMO conjugation to substrate target proteins (sumoylation) in Arabidopsis (Arabidopsis thaliana). siz1 T-DNA insertional mutations (siz1-2 and siz1-3; Miura et al., 2005) cause basal, but not acquired, thermosensitivity that occurs in conjunction with hyperaccumulation of salicylic acid (SA). NahG encodes a salicylate hydroxylase, and expression in siz1-2 seedlings reduces endogenous SA accumulation to that of wild-type levels and further increases thermosensitivity. High temperature induces SUMO1/2 conjugation to peptides in wild type but to a substantially lesser degree in siz1 mutants. However, heat shock-induced expression of genes, including heat shock proteins, ascorbate peroxidase 1 and 2, is similar in siz1 and wild-type seedlings. Together, these results indicate that SIZ1 and, by inference, sumoylation facilitate basal thermotolerance through processes that are SA independent.
Figures
Similar articles
-
Salicylic acid-mediated innate immunity in Arabidopsis is regulated by SIZ1 SUMO E3 ligase.Plant J. 2007 Jan;49(1):79-90. doi: 10.1111/j.1365-313X.2006.02947.x. Epub 2006 Dec 6. Plant J. 2007. PMID: 17163880
-
E3 SUMO ligase SIZ1 splicing variants localize and function according to external conditions.Plant Physiol. 2024 May 31;195(2):1601-1623. doi: 10.1093/plphys/kiae108. Plant Physiol. 2024. PMID: 38497423 Free PMC article.
-
An Arabidopsis SUMO E3 Ligase, SIZ1, Negatively Regulates Photomorphogenesis by Promoting COP1 Activity.PLoS Genet. 2016 Apr 29;12(4):e1006016. doi: 10.1371/journal.pgen.1006016. eCollection 2016 Apr. PLoS Genet. 2016. PMID: 27128446 Free PMC article.
-
SIZ1-mediated SUMOylation during phosphate homeostasis in plants: Looking beyond the tip of the iceberg.Semin Cell Dev Biol. 2018 Feb;74:123-132. doi: 10.1016/j.semcdb.2017.09.016. Epub 2017 Sep 10. Semin Cell Dev Biol. 2018. PMID: 28903074 Review.
-
Regulation of Plant Cellular and Organismal Development by SUMO.Adv Exp Med Biol. 2017;963:227-247. doi: 10.1007/978-3-319-50044-7_14. Adv Exp Med Biol. 2017. PMID: 28197916 Review.
Cited by
-
Arabidopsis small ubiquitin-like modifier paralogs have distinct functions in development and defense.Plant Cell. 2010 Jun;22(6):1998-2016. doi: 10.1105/tpc.109.070961. Epub 2010 Jun 4. Plant Cell. 2010. PMID: 20525853 Free PMC article.
-
Flowering time regulation by the SUMO E3 ligase SIZ1.Plant Signal Behav. 2008 Oct;3(10):891-2. doi: 10.4161/psb.3.10.6513. Plant Signal Behav. 2008. PMID: 19704532 Free PMC article.
-
Regulation of cold signaling by sumoylation of ICE1.Plant Signal Behav. 2008 Jan;3(1):52-3. doi: 10.4161/psb.3.1.4865. Plant Signal Behav. 2008. PMID: 19704769 Free PMC article.
-
Transcriptional memory and response to adverse temperatures in plants.J Zhejiang Univ Sci B. 2021 Oct 15;22(10):791-804. doi: 10.1631/jzus.B2100287. J Zhejiang Univ Sci B. 2021. PMID: 34636184 Free PMC article. Review. English.
-
SUMO and SUMOylation in plants.Mol Cells. 2011 Oct;32(4):305-16. doi: 10.1007/s10059-011-0122-7. Epub 2011 Sep 9. Mol Cells. 2011. PMID: 21912873 Free PMC article. Review.
References
-
- Alfonso M, Yruela I, Almárcegui S, Torrado E, Pérez MA, Picorel R (2001) Unusual tolerance to high temperatures in a new herbicide-resistant D1 mutant from Glycine max (L.) Merr. cell cultures deficient in fatty acid desaturation. Planta 212: 573–582 - PubMed
-
- Baniwal SK, Bharti K, Chan KY, Fauth M, Ganguli A, Kotak S, Mishra SK, Nover L, Port M, Scharf KD, et al (2004) Heat stress response in plants: a complex game with chaperones and more than twenty heat stress transcription factors. J Biosci 29: 471–487 - PubMed
-
- Bernier-Villamor V, Sampson DA, Matunis MJ, Lima CD (2002) Structural basis for E2-mediated SUMO conjugation revealed by a complex between ubiquitin-conjugating enzyme Ubc9 and RanGAP1. Cell 108: 345–356 - PubMed
-
- Berry J, Björkman O (1980) Photosynthetic response and adaptation to temperature in higher plants. Annu Rev Plant Physiol 31: 491–543
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
