

Heterologous expression and molecular and cellular characterization of CaPUB1 encoding a hot pepper U-Box E3 ubiquitin ligase homolog
- PMID: 17041029
- PMCID: PMC1676043
- DOI: 10.1104/pp.106.087965
Heterologous expression and molecular and cellular characterization of CaPUB1 encoding a hot pepper U-Box E3 ubiquitin ligase homolog
Abstract
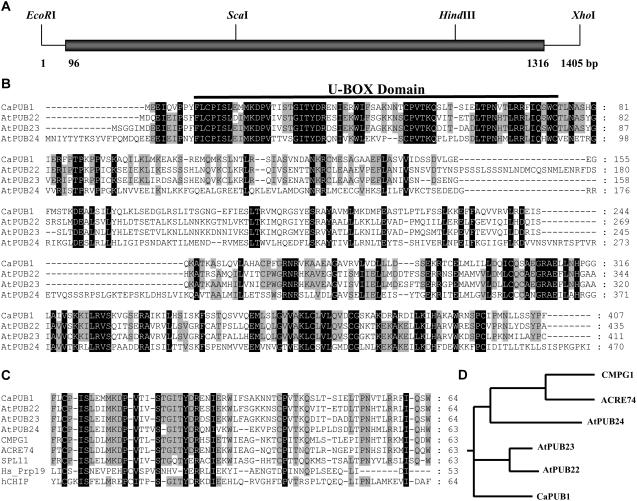
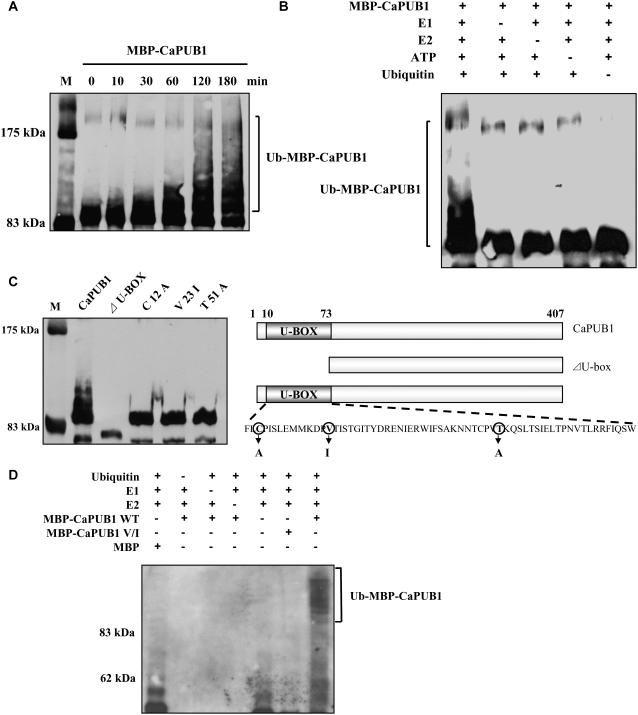
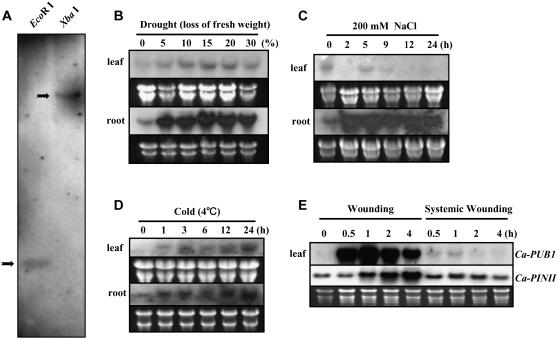
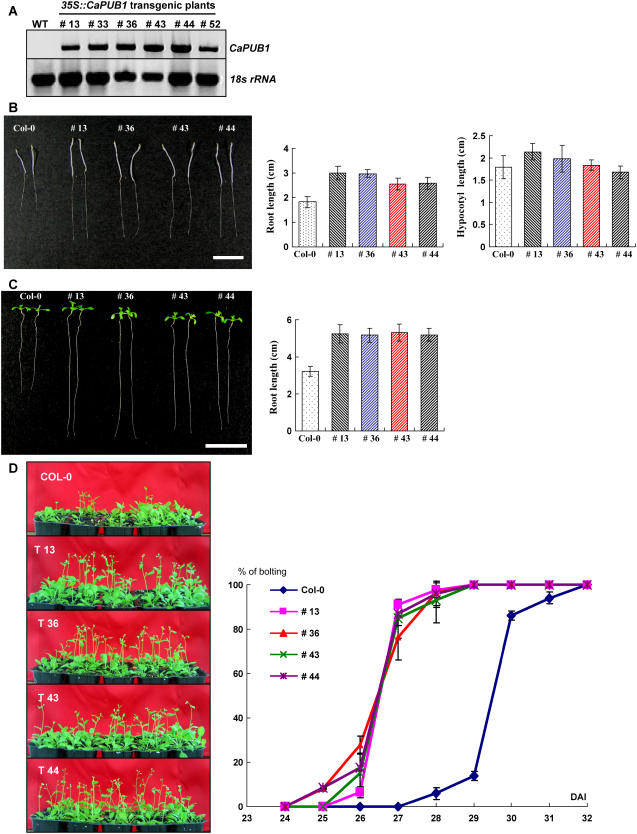
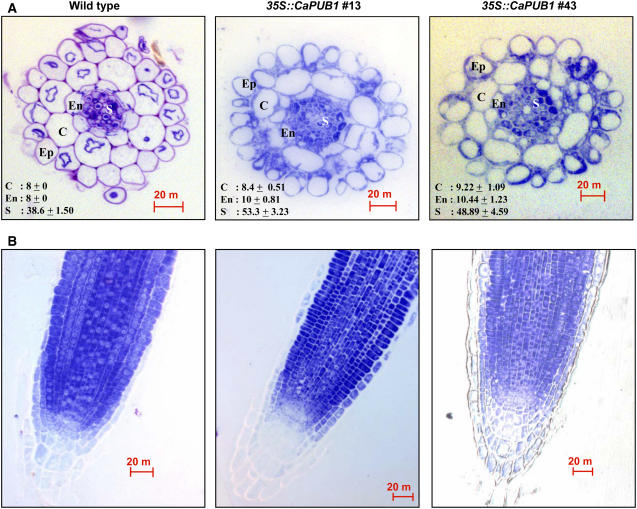
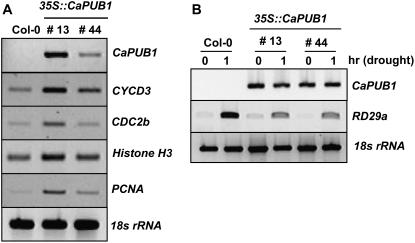
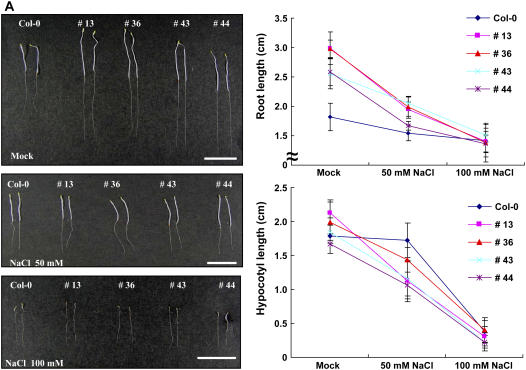
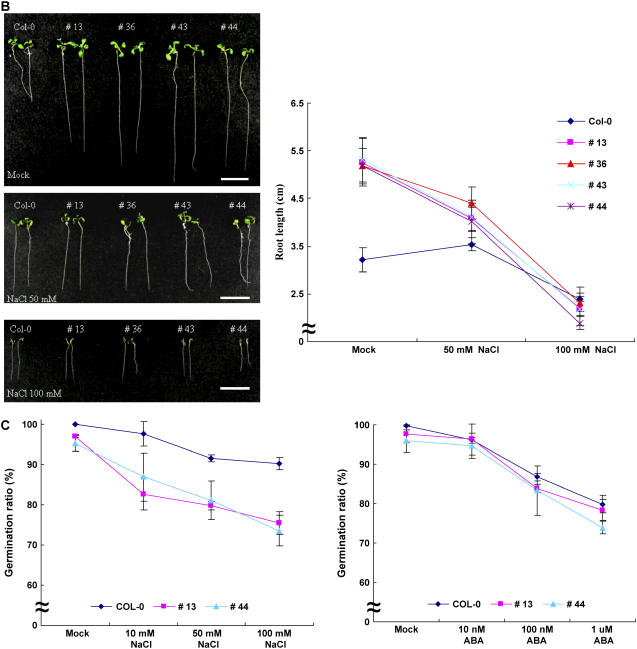
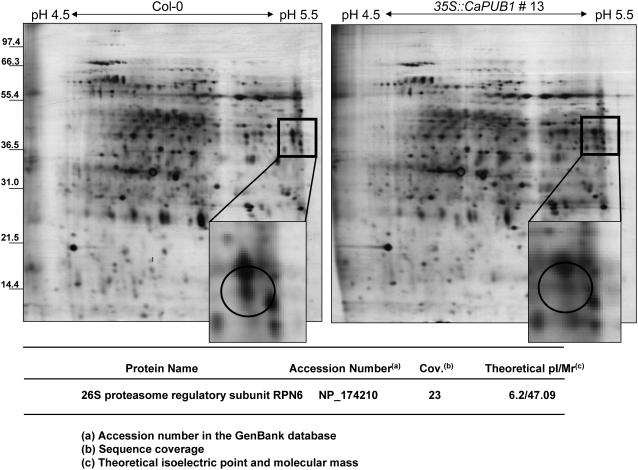
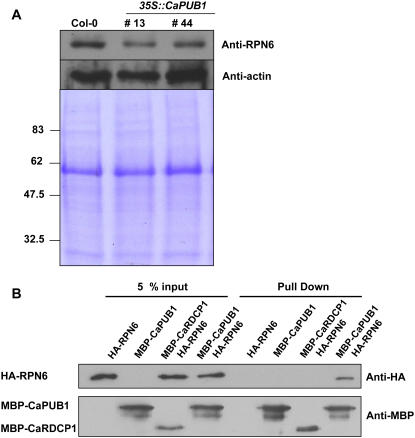
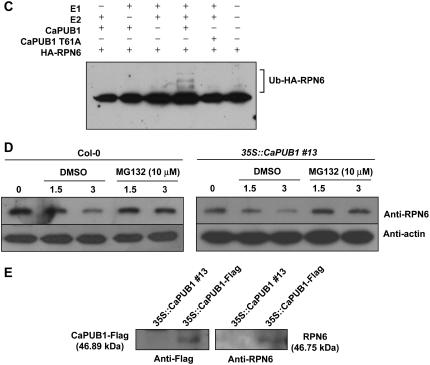
The U-box motif is a conserved domain found in the diverse isoforms of E3 ubiquitin ligase in eukaryotes. From water-stressed hot pepper (Capsicum annuum L. cv Pukang) plants, we isolated C. annuum putative U-box protein 1 (CaPUB1), which encodes a protein containing a single U-box motif in its N-terminal region. In vitro ubiquitination and site-directed mutagenesis assays revealed that CaPUB1 possessed E3 ubiquitin ligase activity and that the U-box motif was indeed essential for its enzyme activity. RNA gel-blot analysis showed that CaPUB1 mRNA was induced rapidly by a broad spectrum of abiotic stresses, including drought, high salinity, cold temperature, and mechanical wounding, but not in response to ethylene, abscisic acid, or a bacterial pathogen, suggesting its role in the early events in the abiotic-related defense response. Because transgenic work was extremely difficult in hot pepper, in this study we overexpressed CaPUB1 in Arabidopsis (Arabidopsis thaliana) to provide cellular information on the function of this gene in the development and plant responses to abiotic stresses. Transgenic Arabidopsis plants that constitutively expressed the CaPUB1 gene under the control of the cauliflower mosaic virus 35S promoter had markedly longer hypocotyls and roots and grew more rapidly than the wild type, leading to an early bolting phenotype. Microscopic analysis showed that 35S::CaPUB1 roots had increased numbers of small-sized cells, resulting in disordered, highly populated cell layers in the cortex, endodermis, and stele. In addition, CaPUB1-overexpressing plants displayed increased sensitivity to water stress and mild salinity. These results indicate that CaPUB1 is functional in Arabidopsis cells, thereby effectively altering cell and tissue growth and also the response to abiotic stresses. Comparative proteomic analysis showed that the level of RPN6 protein, a non-ATPase subunit of the 26S proteasome complex, was significantly reduced in 35SCaPUB1 seedlings as compared to the wild type. Pull-down and ubiquitination assays demonstrated that RPN6 interacted physically with CaPUB1 and was ubiquitinated in a CaPUB1-dependent manner in vitro. Although the physiological function of CaPUB1 is not yet clear, there are several possibilities for its involvement in a subset of physiological responses to counteract dehydration and high-salinity stresses in transgenic Arabidopsis seedlings.
Figures

Similar articles
-
Constitutive expression of abiotic stress-inducible hot pepper CaXTH3, which encodes a xyloglucan endotransglucosylase/hydrolase homolog, improves drought and salt tolerance in transgenic Arabidopsis plants.FEBS Lett. 2006 May 29;580(13):3136-44. doi: 10.1016/j.febslet.2006.04.062. Epub 2006 May 2. FEBS Lett. 2006. PMID: 16684525
-
CaPUB1, a Hot Pepper U-box E3 Ubiquitin Ligase, Confers Enhanced Cold Stress Tolerance and Decreased Drought Stress Tolerance in Transgenic Rice (Oryza sativa L.).Mol Cells. 2016 Mar;39(3):250-7. doi: 10.14348/molcells.2016.2290. Epub 2015 Dec 16. Mol Cells. 2016. PMID: 26674966 Free PMC article.
-
RING Type E3 Ligase CaAIR1 in Pepper Acts in the Regulation of ABA Signaling and Drought Stress Response.Plant Cell Physiol. 2015 Sep;56(9):1808-19. doi: 10.1093/pcp/pcv103. Epub 2015 Jul 13. Plant Cell Physiol. 2015. PMID: 26169196
-
RING E3 ligases: key regulatory elements are involved in abiotic stress responses in plants.BMB Rep. 2017 Aug;50(8):393-400. doi: 10.5483/bmbrep.2017.50.8.128. BMB Rep. 2017. PMID: 28712388 Free PMC article. Review.
-
News from the PUB: plant U-box type E3 ubiquitin ligases.J Exp Bot. 2018 Jan 23;69(3):371-384. doi: 10.1093/jxb/erx411. J Exp Bot. 2018. PMID: 29237060 Review.
Cited by
-
Overexpression of VrUBC1, a Mung Bean E2 Ubiquitin-Conjugating Enzyme, Enhances Osmotic Stress Tolerance in Arabidopsis.PLoS One. 2013 Jun 18;8(6):e66056. doi: 10.1371/journal.pone.0066056. Print 2013. PLoS One. 2013. PMID: 23824688 Free PMC article.
-
Genetic basis of drought tolerance during seed germination in barley.PLoS One. 2018 Nov 2;13(11):e0206682. doi: 10.1371/journal.pone.0206682. eCollection 2018. PLoS One. 2018. PMID: 30388157 Free PMC article.
-
The Arabidopsis RING E3 ubiquitin ligase AtAIRP2 plays combinatory roles with AtAIRP1 in abscisic acid-mediated drought stress responses.Plant Physiol. 2011 Dec;157(4):2240-57. doi: 10.1104/pp.111.185595. Epub 2011 Oct 10. Plant Physiol. 2011. PMID: 21969385 Free PMC article.
-
Altered germination and subcellular localization patterns for PUB44/SAUL1 in response to stress and phytohormone treatments.PLoS One. 2011;6(6):e21321. doi: 10.1371/journal.pone.0021321. Epub 2011 Jun 27. PLoS One. 2011. PMID: 21738636 Free PMC article.
-
Genome-wide survey and expression analysis of the PUB family in Chinese cabbage (Brassica rapa ssp. pekinesis).Mol Genet Genomics. 2015 Dec;290(6):2241-60. doi: 10.1007/s00438-015-1075-x. Epub 2015 Jun 9. Mol Genet Genomics. 2015. PMID: 26054324
References
-
- Azevedo C, Santos-Rosa MJ, Shirasu K (2001) The U-box protein family in plants. Trends Plant Sci 6: 354–358 - PubMed
-
- Baumeister W, Walz J, Zuhl F, Seemuller E (1998) The proteasome: paradigm of a self-compartmentalizing protease. Cell 92: 367–380 - PubMed
-
- Boyer JS (1982) Plant productivity and environment. Science 218: 443–448 - PubMed
-
- Bray EA (1997) Plant responses to water deficit. Trends Plant Sci 2: 48–54
-
- Cho SK, Kim JE, Park J-A, Eom TJ, Kim WT (2006) Constitutive expression of abiotic stress-inducible hot pepper CaXTH3, which encodes a xyloglucan endotransglucosylase/hydrolase homolog, improves drought and salt tolerance in transgenic Arabidopsis plants. FEBS Lett 580: 3136–3144 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
