

NAD(P)H oscillates in pollen tubes and is correlated with tip growth
- PMID: 17041030
- PMCID: PMC1676060
- DOI: 10.1104/pp.106.087882
NAD(P)H oscillates in pollen tubes and is correlated with tip growth
Abstract
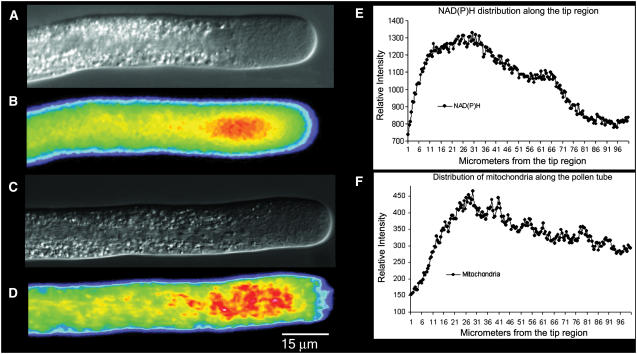
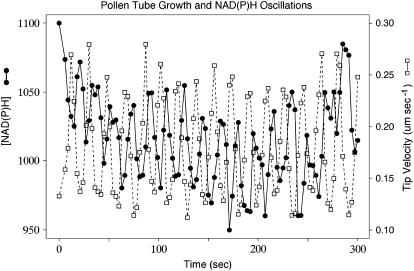
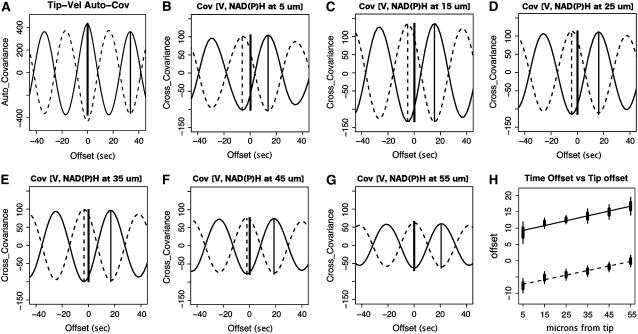
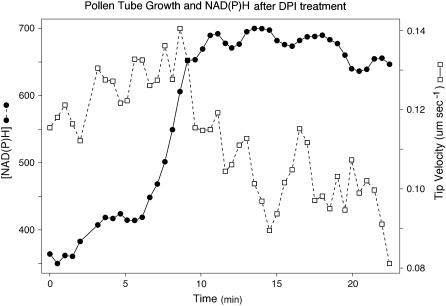
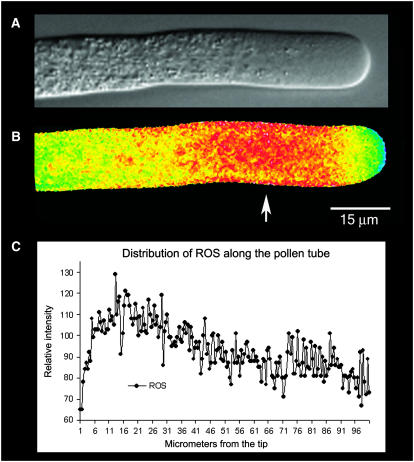
The location and changes in NAD(P)H have been monitored during oscillatory growth in pollen tubes of lily (Lilium formosanum) using the endogenous fluorescence of the reduced coenzyme (excitation, 360 nm; emission, >400 nm). The strongest signal resides 20 to 40 microm behind the apex where mitochondria (stained with Mitotracker Green) accumulate. Measurements at 3-s intervals reveal that NAD(P)H-dependent fluorescence oscillates during oscillatory growth. Cross-correlation analysis indicates that the peaks follow growth maxima by 7 to 11 s or 77 degrees to 116 degrees, whereas the troughs anticipate growth maxima by 5 to 10 s or 54 degrees to 107 degrees. We have focused on the troughs because they anticipate growth and are as strongly correlated with growth as the peaks. Analysis of the signal in 10-microm increments along the length of the tube indicates that the troughs are most advanced in the extreme apex. However, this signal moves basipetally as a wave, being in phase with growth rate oscillations at 50 to 60 microm from the apex. We suggest that the changes in fluorescence are due to an oscillation between the reduced (peaks) and oxidized (troughs) states of the coenzyme and that an increase in the oxidized state [NAD(P)(+)] may be coupled to the synthesis of ATP. We also show that diphenyleneiodonium, an inhibitor of NAD(P)H dehydrogenases, causes an increase in fluorescence and a decrease in tube growth. Finally, staining with 5-(and-6)-chloromethyl-2',7'-dichlorohydrofluorescein acetate indicates that reactive oxygen species are most abundant in the region where mitochondria accumulate and where NAD(P)H fluorescence is maximal.
Figures

References
-
- Baker CJ, Deahl K, Domek J, Orlandi EW (1998) Oxygen metabolism in plant/bacteria interactions: effect of DPI on the pseudo-NAD(P)H oxidase activity of peroxidase. Biochem Biophys Res Commun 252: 461–464 - PubMed
-
- Blanchoin L, Pollard TD (1999) Mechanism of interaction of Acanthamoeba actophorin (ADF/Cofilin) with actin filaments. J Biol Chem 274: 15538–15546 - PubMed
-
- Blinova K, Carroll S, Bose S, Smirnov AV, Harvey JJ, Knutson JR, Balaban RS (2005) Distribution of mitochondrial NADH fluorescence lifetimes: steady-state kinetics of matrix NADH interactions. Biochemistry 44: 2585–2594 - PubMed
-
- Brachmanski M, Gebhard MM, Nobiling R (2004) Separation of fluorescence signals from Ca2+ and NADH during cardioplegic arrest and cardiac ischemia. Cell Calcium 35: 381–391 - PubMed
-
- Brillinger DR (1975) Time Series: Data Analysis and Theory. Holt-Rinehart, New York
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
