

Review
doi: 10.1128/JVI.00919-06.
Nonsegmented negative-strand viruses as vaccine vectors
Affiliations
- PMID: 17041210
- PMCID: PMC1641758
- DOI: 10.1128/JVI.00919-06
Item in Clipboard
Review
Nonsegmented negative-strand viruses as vaccine vectors
J Virol.
2006 Nov.
No abstract available
Figures

Features of NNSV that make them attractive as viral vectors.
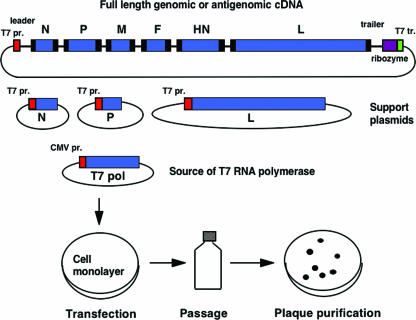
Recovery of a complete, infectious NNSV from cDNA. A plasmid encoding the complete antigenomic RNA of a parainfluenza virus is shown at the top. The extragenic 3′ leader and 5′ trailer regions are shown, as are the six viral genes: N, nucleoprotein; P, phosphoprotein, M, matrix protein, F, fusion protein, HN, hemagglutinin-neuraminidase, and L, large polymerase subunit. The GS and GE transcription signals are shown as black boxes at the beginning and at the end of each gene, respectively. The antigenomic cDNA is flanked to the left by a promoter for T7 RNA polymerase (T7 pr.) and on the right by a self-cleaving ribozyme and T7 terminator (T7 tr.). Three other T7 expression plasmids encode the N, P, and L support proteins needed to reconstitute a biologically active nucleocapsid. The T7 RNA polymerase is supplied from a cotransfected eukaryotic expression plasmid bearing a cytomegalovirus promoter (CMV pr.). The plasmids are transfected or electroporated into a cell monolayer in which the plasmid-expressed viral RNA and protein components assemble into a functional nucleocapsid and launch a productive infection. Recovered virus is amplified by passage and can be plaque purified.
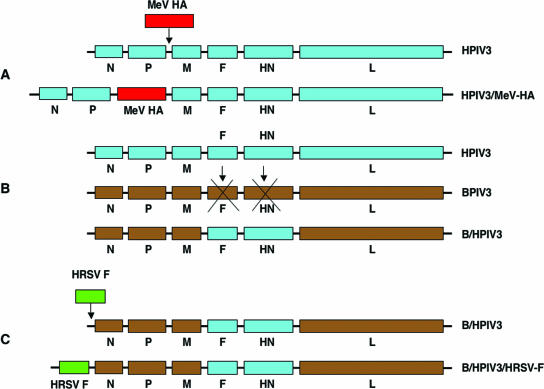
Three strategies for designing NNSV vectors. In panel A, a complete virus (in this case, HPIV3) is modified by insertion of a transcription cassette encoding a foreign antigen (measles virus [MeV] HA glycoprotein). In panel B, a NNSV (BPIV3) is modified by deleting its surface glycoprotein genes (F and HN) and replacing them with those (F and HN) from the target pathogen (HPIV3), resulting in an antigenic chimeric virus (B/HPIV3). In panel C, an antigenic chimeric virus (B/HPIV3) is modified by insertion of a transcription cassette encoding a foreign glycoprotein (HRSV F). In each case (A to C), the coding sequence of the foreign glycoprotein(s) must be under the control of GS and GE signals that are compatible with the vector backbone.
Examples of the incorporation of foreign glycoproteins into NNSV vector particles involving antigenic chimeric viruses (B and C) and foreign glycoproteins expressed from an added gene (D and E). The F and HN glycoproteins of HPIV3 (A, black) were deleted and replaced with the F and HN glycoproteins of HPIV1 (red), yielding an antigenic chimeric virus bearing the surface antigens of HPIV1 (B). A comparable derivative of HPIV3 bearing the surface antigens of HPIV2 (purple) was viable only if the HPIV2 F and HN proteins were modified to contain the cytoplasmic tails of the HPIV3 F and HN proteins, respectively (C). The expression of Ebola virus GP (green) from an added gene in HPIV3 resulted the incorporation of GP into the HPIV3 vector particle (D), whereas the incorporation of the avian influenza HA protein (FLU, orange) expressed from an added gene in NDV (blue) depended on replacing its cytoplasmic and transmembrane domains with those of the NDV F protein (E). ecto, ectodomain; tm, transmembrane domain; cyto, cytoplasmic domain.
Comment in
-
Caution about Newcastle disease virus-based live attenuated vaccine.J Virol. 2008 Jul;82(13):6782. doi: 10.1128/JVI.00370-08. J Virol. 2008. PMID: 18544791 Free PMC article. No abstract available.
References
-
- Bartlett, E. J., E. Amaro-Carambot, S. R. Surman, J. T. Newman, P. L. Collins, B. R. Murphy, and M. H. Skiadopoulos. 2005. Human parainfluenza virus type I (HPIV1) vaccine candidates designed by reverse genetics are attenuated and efficacious in African green monkeys. Vaccine 23:4631-4646. - PubMed
-
- Buchholz, U. J., S. Biacchesi, Q. N. Pham, K. C. Tran, L. Yang, C. L. Luongo, M. H. Skiadopoulos, B. R. Murphy, and P. L. Collins. 2005. Deletion of M2 gene open reading frames 1 and 2 of human metapneumovirus: effects on RNA synthesis, attenuation, and immunogenicity. J. Virol. 79:6588-6597. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
