

Abnormal skin, limb and craniofacial morphogenesis in mice deficient for interferon regulatory factor 6 (Irf6)
- PMID: 17041601
- PMCID: PMC2082114
- DOI: 10.1038/ng1903
Abnormal skin, limb and craniofacial morphogenesis in mice deficient for interferon regulatory factor 6 (Irf6)
Abstract
Transcription factor paralogs may share a common role in staged or overlapping expression in specific tissues, as in the Hox family. In other cases, family members have distinct roles in a range of embryologic, differentiation or response pathways (as in the Tbx and Pax families). For the interferon regulatory factor (IRF) family of transcription factors, mice deficient in Irf1, Irf2, Irf3, Irf4, Irf5, Irf7, Irf8 or Irf9 have defects in the immune response but show no embryologic abnormalities. Mice deficient for Irf6 have not been reported, but in humans, mutations in IRF6 cause two mendelian orofacial clefting syndromes, and genetic variation in IRF6 confers risk for isolated cleft lip and palate. Here we report that mice deficient for Irf6 have abnormal skin, limb and craniofacial development. Histological and gene expression analyses indicate that the primary defect is in keratinocyte differentiation and proliferation. This study describes a new role for an IRF family member in epidermal development.
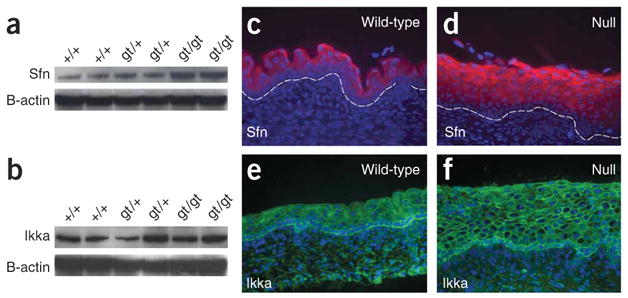
Figures
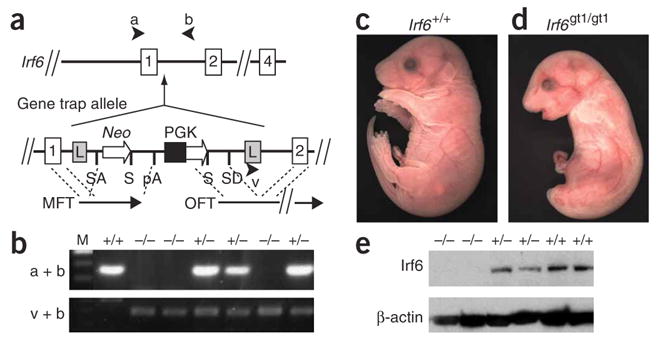
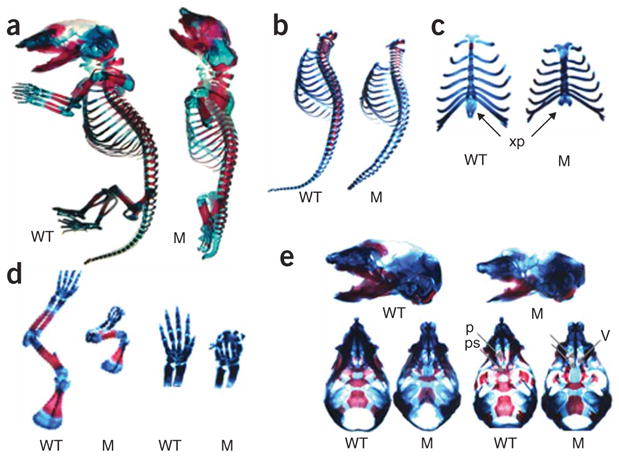
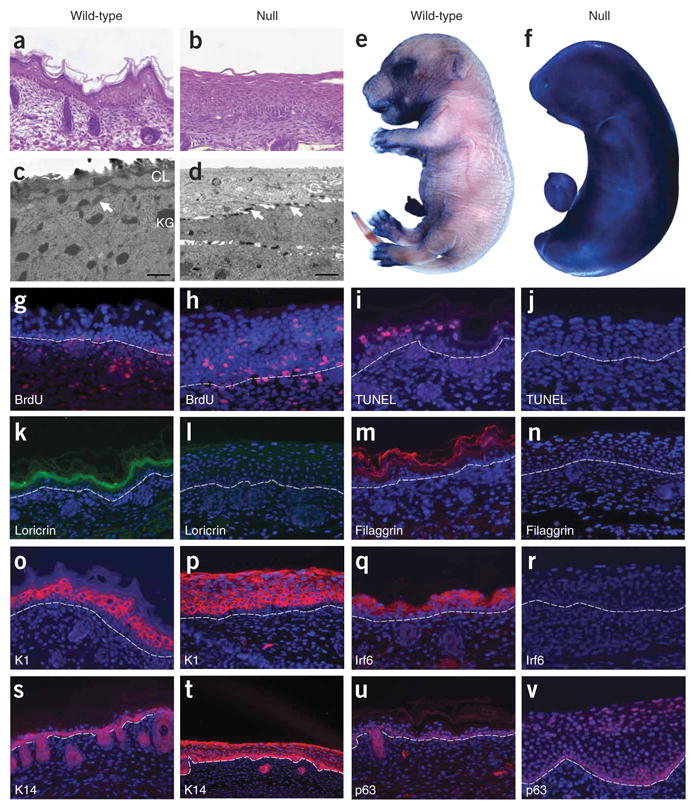
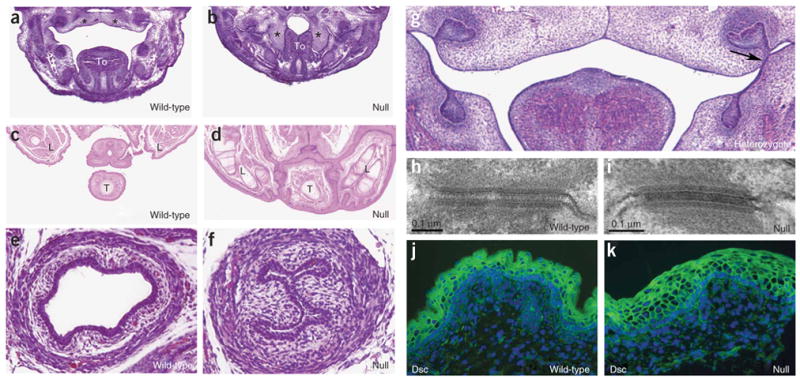
References
-
- Matsuyama T, et al. Targeted disruption of IRF-1 or IRF-2 results in abnormal type I IFN gene induction and aberrant lymphocyte development. Cell. 1993;75:83–97. - PubMed
-
- Sato M, et al. Distinct and essential roles of transcription factors IRF-3 and IRF-7 in response to viruses for IFN-alpha/beta gene induction. Immunity. 2000;13:539–548. - PubMed
-
- Mittrucker HW, et al. Requirement for the transcription factor LSIRF/IRF4 for mature B and T lymphocyte function. Science. 1997;275:540–543. - PubMed
-
- Takaoka A, et al. Integral role of IRF-5 in the gene induction programme activated by Toll-like receptors. Nature. 2005;434:243–249. - PubMed
-
- Honda K, et al. IRF-7 is the master regulator of type-I interferon-dependent immune responses. Nature. 2005;434:772–777. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
