

Alterations of metabolic activity in human osteoarthritic osteoblasts by lipid peroxidation end product 4-hydroxynonenal
- PMID: 17042956
- PMCID: PMC1794501
- DOI: 10.1186/ar2066
Alterations of metabolic activity in human osteoarthritic osteoblasts by lipid peroxidation end product 4-hydroxynonenal
Abstract
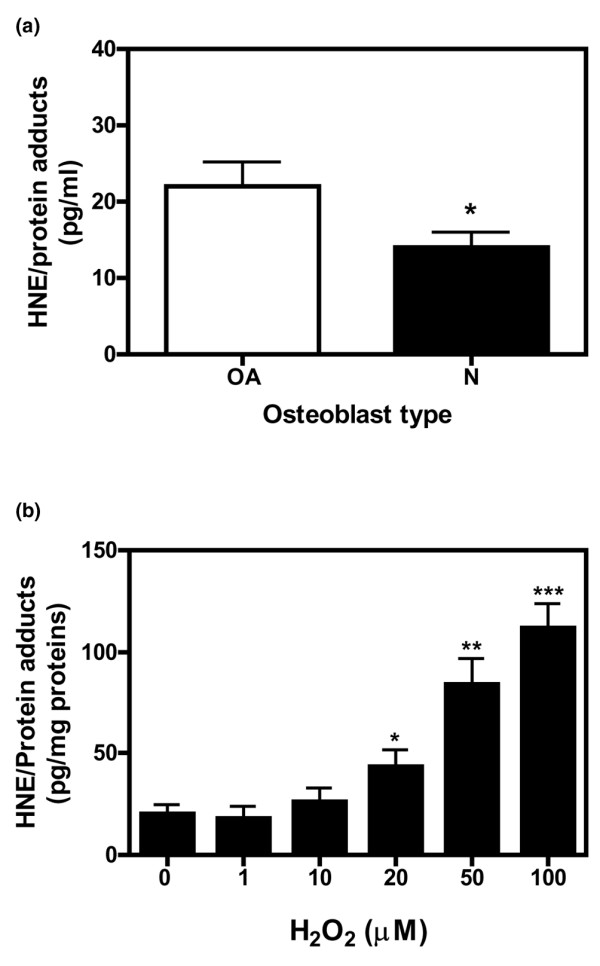
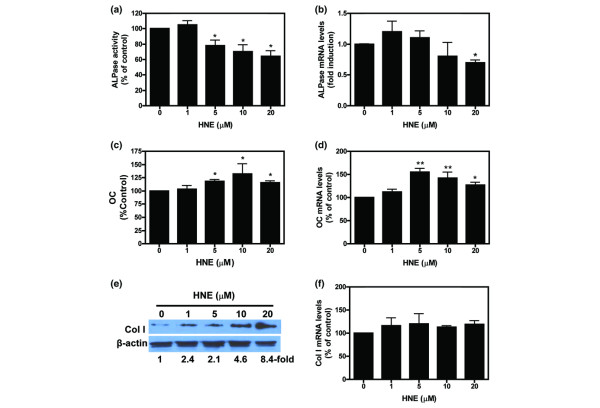
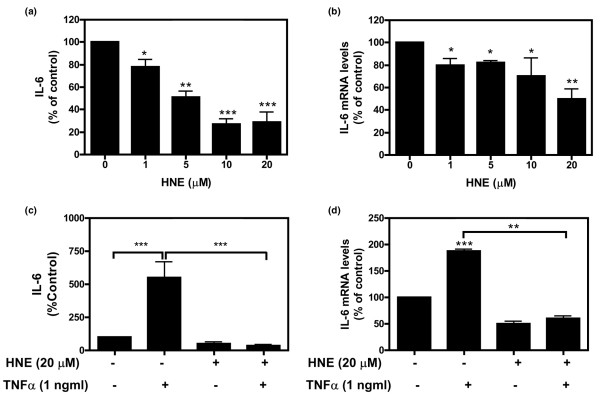
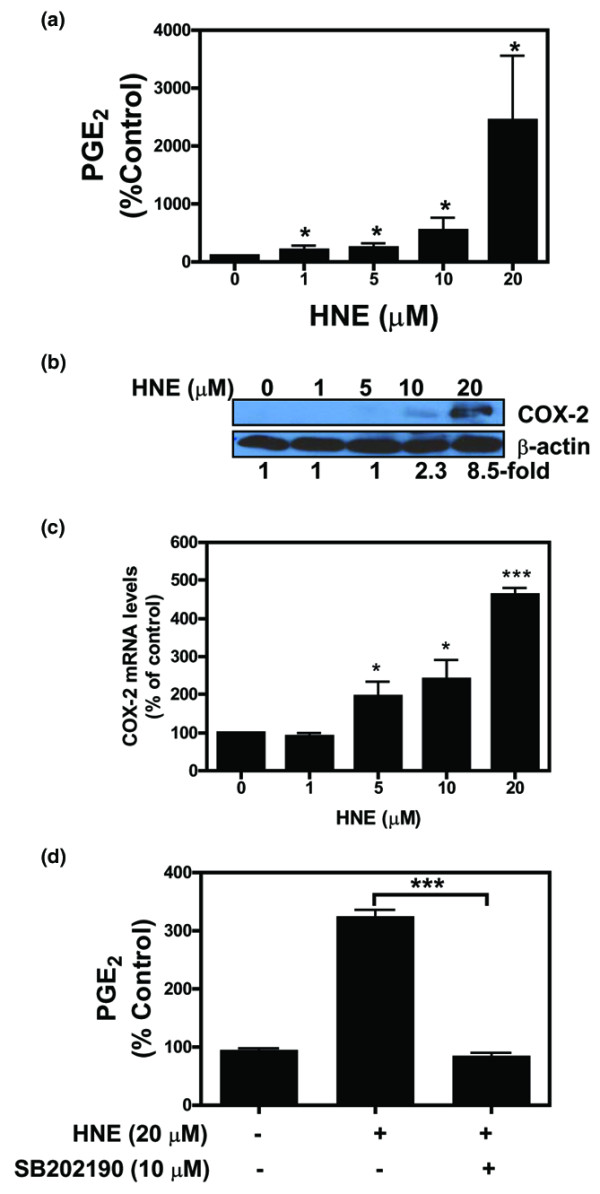
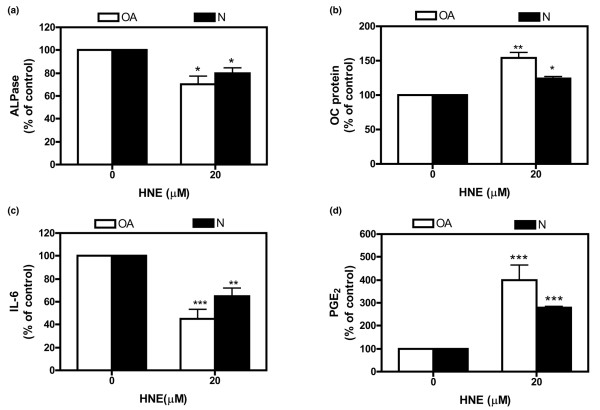
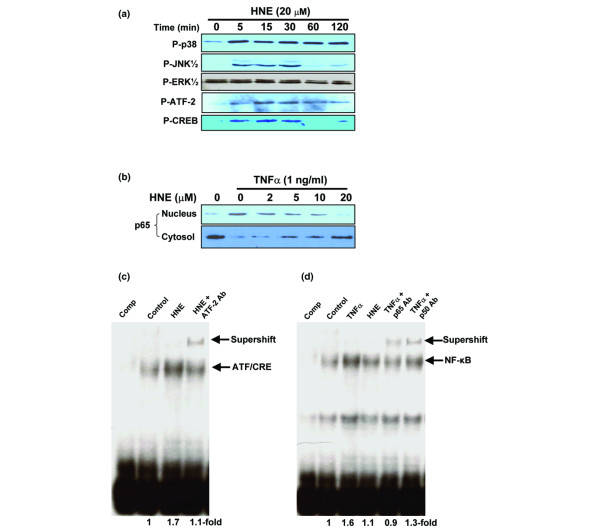
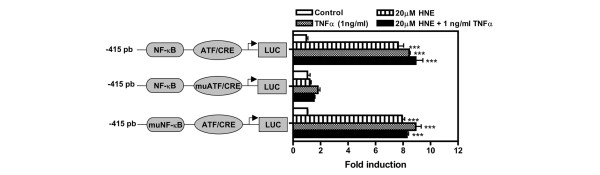
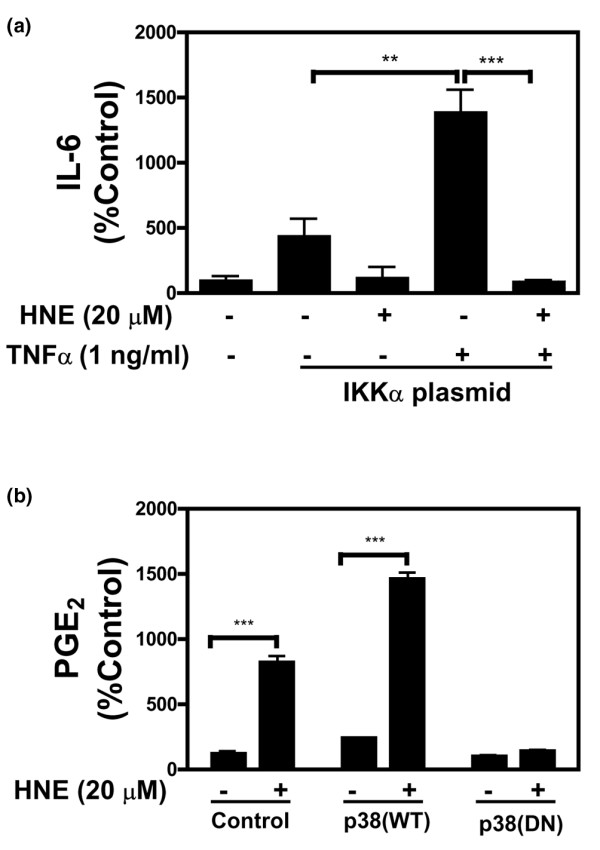
4-Hydroxynonenal (HNE), a lipid peroxidation end product, is produced abundantly in osteoarthritic (OA) articular tissues, but its role in bone metabolism is ill-defined. In this study, we tested the hypothesis that alterations in OA osteoblast metabolism are attributed, in part, to increased levels of HNE. Our data showed that HNE/protein adduct levels were higher in OA osteoblasts compared to normal and when OA osteoblasts were treated with H2O2. Investigating osteoblast markers, we found that HNE increased osteocalcin and type I collagen synthesis but inhibited alkaline phosphatase activity. We next examined the effects of HNE on the signaling pathways controlling cyclooxygenase-2 (COX-2) and interleukin-6 (IL-6) expression in view of their putative role in OA pathophysiology. HNE dose-dependently decreased basal and tumour necrosis factor-alpha (TNF-alpha)-induced IL-6 expression while inducing COX-2 expression and prostaglandin E2 (PGE2) release. In a similar pattern, HNE induces changes in osteoblast markers as well as PGE2 and IL-6 release in normal osteoblasts. Upon examination of signaling pathways involved in PGE2 and IL-6 production, we found that HNE-induced PGE2 release was abrogated by SB202190, a p38 mitogen-activated protein kinase (MAPK) inhibitor. Overexpression of p38 MAPK enhanced HNE-induced PGE2 release. In this connection, HNE markedly increased the phosphorylation of p38 MAPK, JNK2, and transcription factors (CREB-1, ATF-2) with a concomitant increase in the DNA-binding activity of CRE/ATF. Transfection experiments with a human COX-2 promoter construct revealed that the CRE element (-58/-53 bp) was essential for HNE-induced COX-2 promoter activity. However, HNE inhibited the phosphorylation of IkappaBalpha and subsequently the DNA-binding activity of nuclear factor-kappaB. Overexpression of IKKalpha increased TNF-alpha-induced IL-6 production. This induction was inhibited when TNF-alpha was combined with HNE. These findings suggest that HNE may exert multiple effects on human OA osteoblasts by selective activation of signal transduction pathways and alteration of osteoblastic phenotype expression and pro-inflammatory mediator production.
Figures
References
-
- Page S, Fischer C, Baumgartner B, Haas M, Kreusel U, Loidl G, Hayn M, Ziegler-Heitbrock HW, Neumeier D, Brand K. 4-Hydroxynonenal prevents NF-kappaB activation and tumor necrosis factor expression by inhibiting IkappaB phosphorylation and subsequent proteolysis. J Biol Chem. 1999;274:11611–11618. doi: 10.1074/jbc.274.17.11611. - DOI - PubMed
-
- Uchida K, Shiraishi M, Naito Y, Torii Y, Nakamura Y, Osawa T. Activation of stress signaling pathways by the end product of lipid peroxidation. 4-hydroxy-2-nonenal is a potential inducer of intracellular peroxide production. J Biol Chem. 1999;274:2234–2242. doi: 10.1074/jbc.274.4.2234. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
