

In vivo evidence that 5-HT(2C) receptors inhibit 5-HT neuronal activity via a GABAergic mechanism
- PMID: 17043669
- PMCID: PMC2014685
- DOI: 10.1038/sj.bjp.0706935
In vivo evidence that 5-HT(2C) receptors inhibit 5-HT neuronal activity via a GABAergic mechanism
Abstract
Background and purpose: Recent evidence suggests that 5-HT(2C) receptor activation may inhibit midbrain 5-HT neurones by activating neighbouring GABA neurones. This hypothesis was tested using the putative selective 5-HT(2C) receptor agonist, WAY 161503.
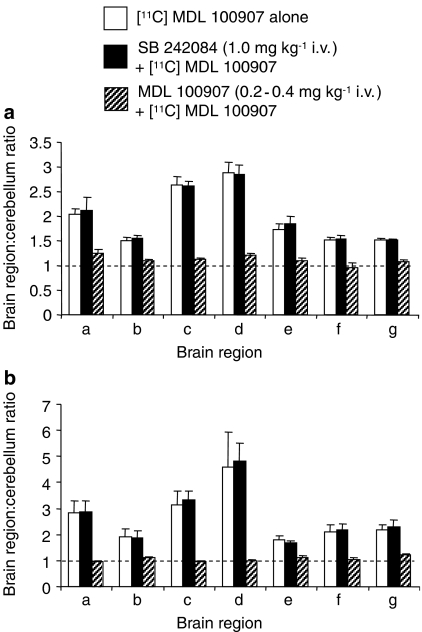
Experimental approach: The effect of WAY 161503 on 5-HT cell firing in the dorsal raphe nucleus (DRN) was investigated in anaesthetised rats using single unit extracellular recordings. The effect of WAY 161503 on DRN GABA neurones was investigated using double label immunohistochemical measurements of Fos, glutamate decarboxylase (GAD) and 5-HT(2C) receptors. Finally, drug occupancy at 5-HT(2A) receptors was investigated using rat positron emission tomography and ex vivo binding studies with the 5-HT(2A) receptor radioligand [(11)C]MDL 100907.
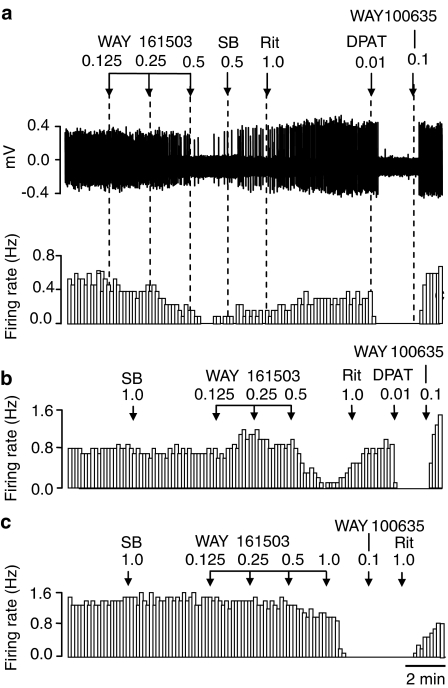
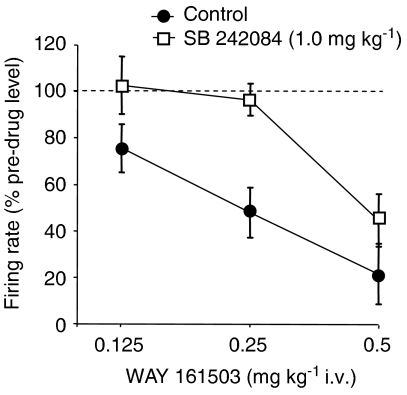
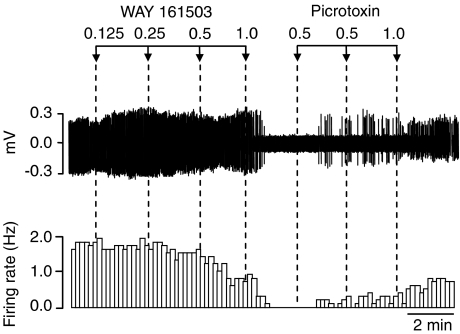
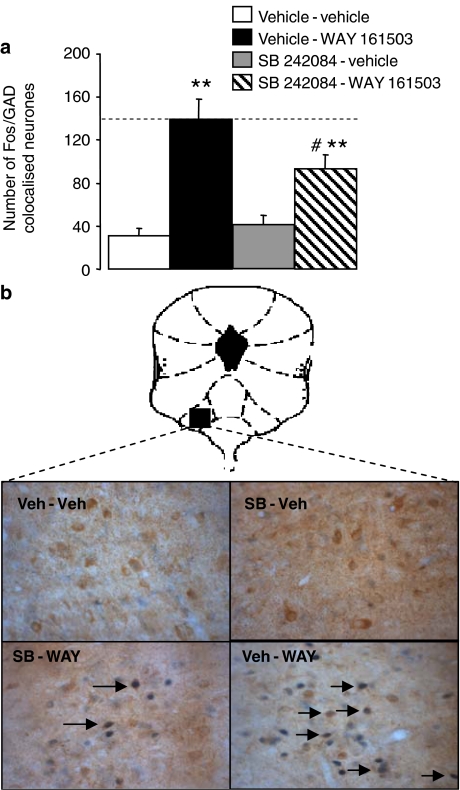
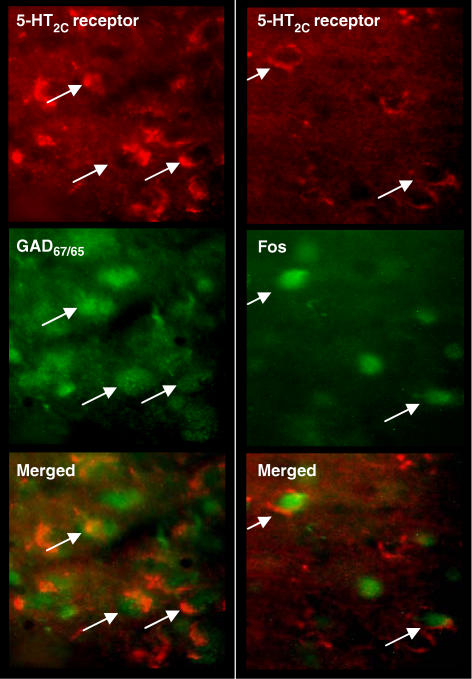
Key results: WAY 161503 caused a dose-related inhibition of 5-HT cell firing which was reversed by the 5-HT(2) receptor antagonist ritanserin and the 5-HT(2C) receptor antagonist SB 242084 but not by the 5-HT(1A) receptor antagonist WAY 100635. SB 242084 pretreatment also prevented the response to WAY 161503. The blocking effects of SB 242084 likely involved 5-HT(2C) receptors because the drug did not demonstrate 5-HT(2A) receptor occupancy in vivo or ex vivo. The inhibition of 5-HT cell firing induced by WAY 161503 was partially reversed by the GABA(A) receptor antagonist picrotoxin. Also, WAY 161503 increased Fos expression in GAD positive DRN neurones and DRN GAD positive neurones expressed 5-HT(2C) receptor immunoreactivity.
Conclusions and implications: These findings indicate that WAY 161503 inhibits 5-HT cell firing in the DRN in vivo, and support a mechanism involving 5-HT(2C) receptor-mediated activation of DRN GABA neurones.
Figures
References
-
- Aghajanian GK, Foote WE, Sheard MH. Action of psychotogenic drugs on single midbrain raphe neurons. J Pharmacol Exp Ther. 1970;171:178–187. - PubMed
-
- Allers KA, Sharp T. Neurochemical and anatomical identification of fast- and slow-firing neurones in the rat dorsal raphe nucleus using juxtacellular labelling methods in vivo. Neuroscience. 2003;122:193–204. - PubMed
-
- Barnes NM, Sharp T. A review of central 5-HT receptors and their function. Neuropharmacology. 1999;38:1083–1152. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
