

Tissue transglutaminase crosslinks ataxin-1: possible role in SCA1 pathogenesis
- PMID: 17045396
- PMCID: PMC2117902
- DOI: 10.1016/j.neulet.2006.08.003
Tissue transglutaminase crosslinks ataxin-1: possible role in SCA1 pathogenesis
Abstract
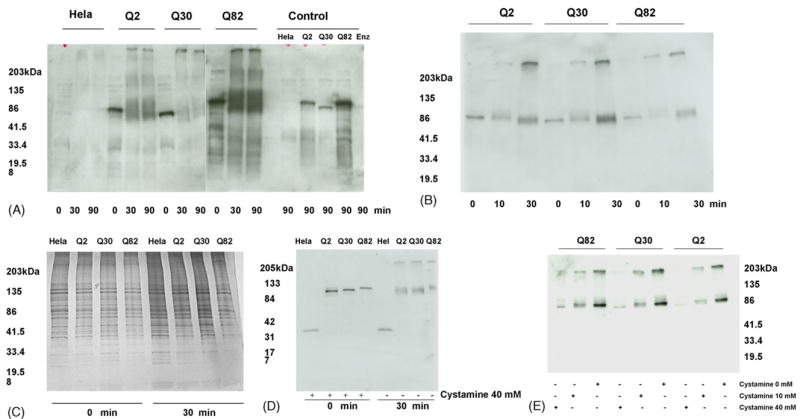
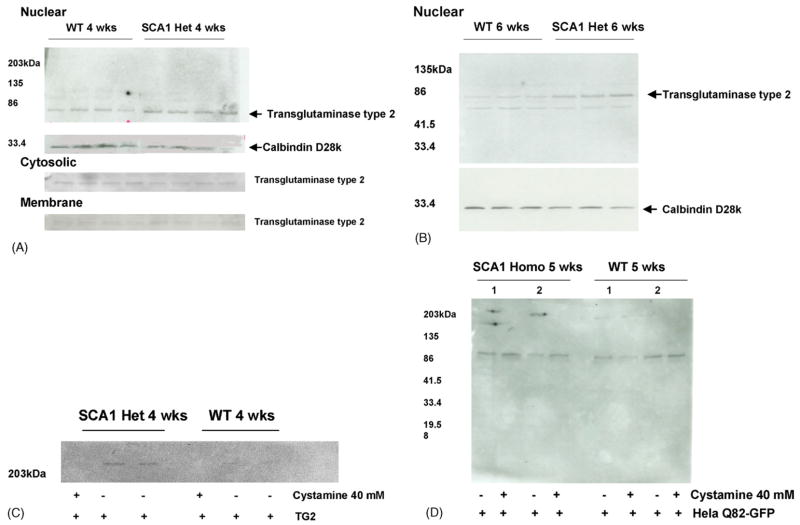
Transglutaminase type 2 (TG2) has recently been implicated in crosslinking of mutant huntingtin protein into aggregates. Here we show that TG2 also crosslinks spinocerebellar ataxia-1 (SCA1) gene product ataxin-1. HeLa cell lysates expressing GFP tagged ataxin-1 with 2, 30 or 82 glutamines showed covalent crosslinking of ataxin-1 when incubated with exogenously added TG2. This crosslinking was inhibited by TG2 inhibitor cystamine. SCA1 transgenic mice which overexpress the mutant ataxin-1 in cerebellar Purkinje cells showed elevated nuclear TG2 in the absence of ataxin-1 nuclear aggregates. The addition of purified TG2 to the nuclear extracts or addition of SCA1 nuclear TG2 to GFP-Q82 HeLa cell lysates resulted in the formation of insoluble aggregates. These data indicate that ataxin-1 is a substrate of TG2. Further, in SCA1 TG2 may translocate to the nucleus in response to nuclear accumulation of mutant ataxin-1 at early stages of the disease.
Figures
References
-
- Aeschlimann D, Tomazy V. Protein crosslinking in assembly and remodeling of extracellular matrices: the role of transglutaminases. Connect Tissue Res. 2000;41:1–27. - PubMed
-
- Bailey CD, Tucholski J, Johnson GV. Transglutaminases in neurodegenerative disorders. Prog Exp Tumor Res. 2005;38:139–157. - PubMed
-
- Burright EN, Clark HB, Servadio A, Matilla T, Feddersen RM, Yunis WS, Duvick LA, Zoghbi HY, Orr HT. SCA1 transgenic mice: a model for neurodegeneration caused by an expanded CAG trinucleotide repeat. Cell. 1995;82:937–948. - PubMed
-
- Gatchel JR, Zoghbi HY. Diseases of unstable repeat expansion: mechanisms and common principles. Nat Rev Genet. 2005;6:743–755. - PubMed
-
- Gentile V, Sepe C, Calvani M, Melone MA, Cotrufo R, Cooper AJ, Blass JP, Peluso G. Tissue transglutaminase-catalyzed formation of high-molecular-weight aggregates in vitro is favored with long polyglutamine domains: a possible mechanism contributing to CAG-triplet diseases. Arch Biochem Biophys. 1998;352:314–321. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
