

Expression analysis of primary mouse megakaryocyte differentiation and its application in identifying stage-specific molecular markers and a novel transcriptional target of NF-E2
- PMID: 17047147
- PMCID: PMC1794061
- DOI: 10.1182/blood-2006-08-038901
Expression analysis of primary mouse megakaryocyte differentiation and its application in identifying stage-specific molecular markers and a novel transcriptional target of NF-E2
Abstract
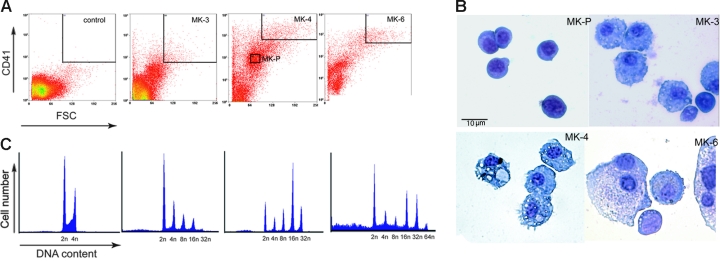
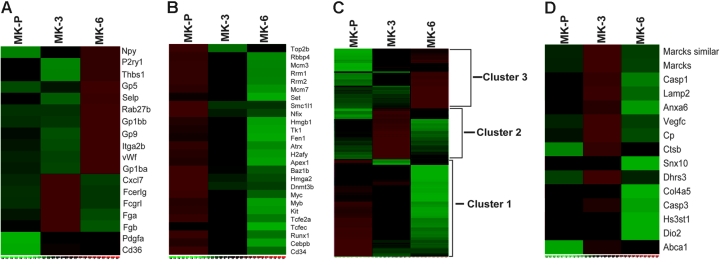
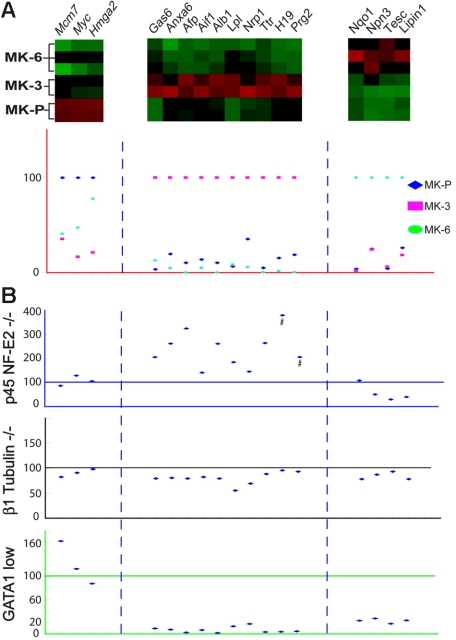
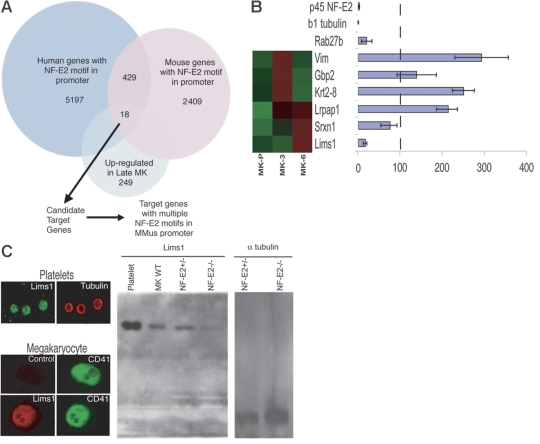
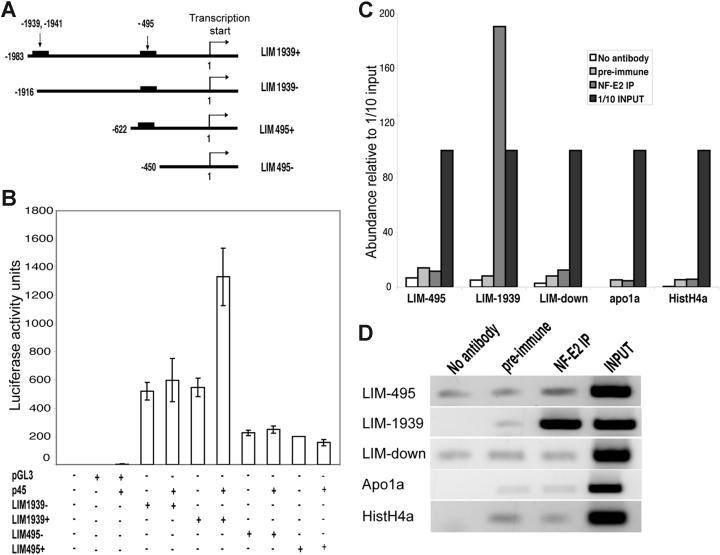
Megakaryocyte (MK) differentiation is well described in morphologic terms but its molecular counterparts and the basis for platelet release are incompletely understood. We profiled mRNA expression in populations of primary mouse MKs representing successive differentiation stages. Genes associated with DNA replication are highly expressed in young MKs, in parallel with endomitosis. Intermediate stages are characterized by disproportionate expression of genes associated with the cytoskeleton, cell migration, and G-protein signaling, whereas terminally mature MKs accumulate hemostatic factors, including many membrane proteins. We used these expression profiles to extract a reliable panel of molecular markers for MKs of early, intermediate, or advanced differentiation and establish the value of this marker panel using mouse models of defective thrombopoiesis resulting from absence of GATA1, NF-E2, or tubulin beta1. Computational analysis of the promoters of late-expressed MK genes identified new candidate targets for NF-E2, a critical transcriptional regulator of platelet release. One such gene encodes the kinase adaptor protein LIMS1/PINCH1, which is highly expressed in MKs and platelets and significantly reduced in NF-E2-deficient cells. Transactivation studies and chromatin immunoprecipitation implicate Lims1 as a direct target of NF-E2 regulation. Attribution of stage-specific genes, in combination with various applications, thus constitutes a powerful way to study MK differentiation and platelet biogenesis.
Figures
References
-
- Ebbe S, Stohlman FJ, Overcash J, Donovan J, Howard D. Megakaryocyte size in thrombocytopenic and normal rats. Blood. 1968;32:383–392. - PubMed
-
- Odell TT, Jackson CW. Polyploidy and maturation of rat megakaryocytes. Blood. 1968;32:102–111. - PubMed
-
- Zucker-Franklin D. Megakaryocytes and platelets. In: Zucker-Franklin D, Greaves MF, Grossi CE, Marmont AM, editors. Atlas of Blood Cells: Function and Pathology. 2nd ed. Vol 1. Philadelphia, PA: Lea & Febiger; 1989. pp. 623–693.
-
- Radley JM, Haller CJ. The demarcation membrane system of the megakaryocyte: a misnomer? Blood. 1982;60:213–219. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
