

Three strategically placed hydrogen-bonding residues convert a proton pump into a sensory receptor
- PMID: 17050685
- PMCID: PMC1637548
- DOI: 10.1073/pnas.0607467103
Three strategically placed hydrogen-bonding residues convert a proton pump into a sensory receptor
Abstract
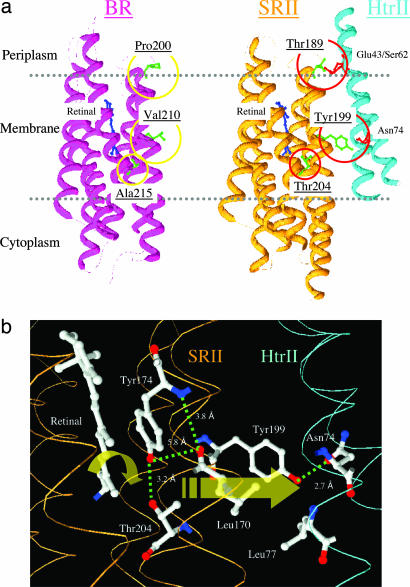
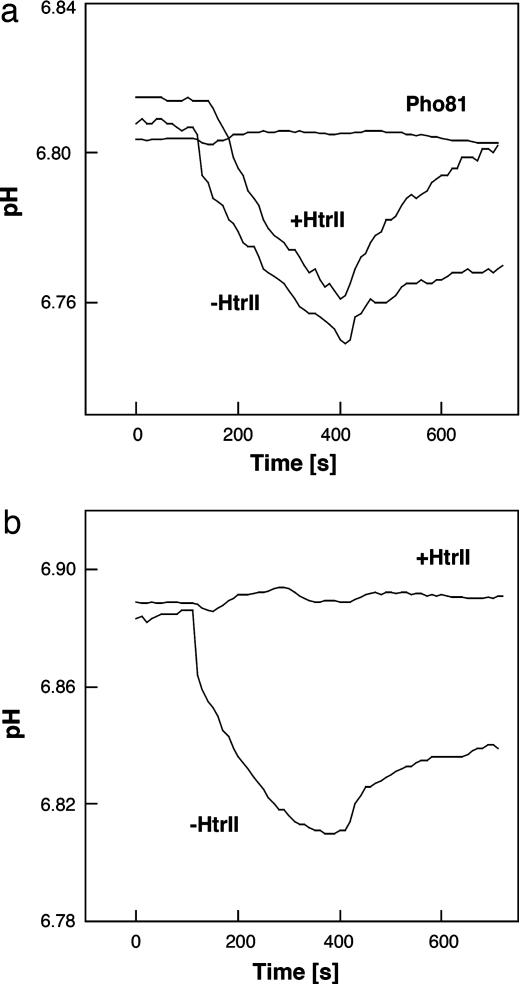
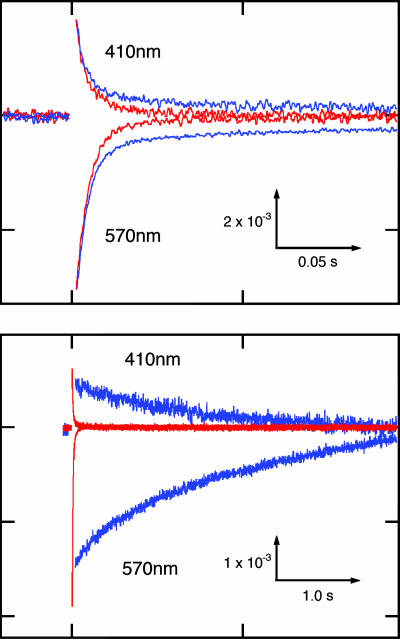
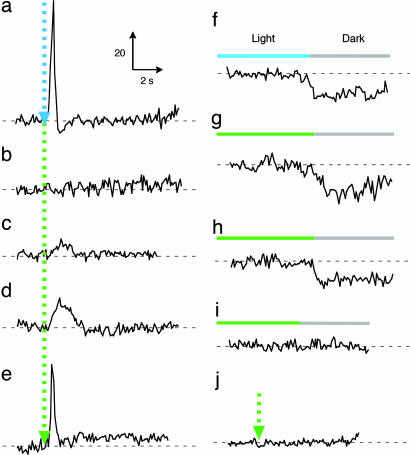
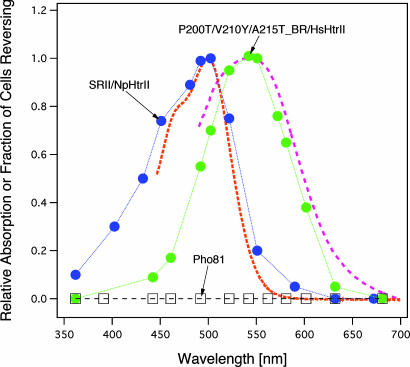
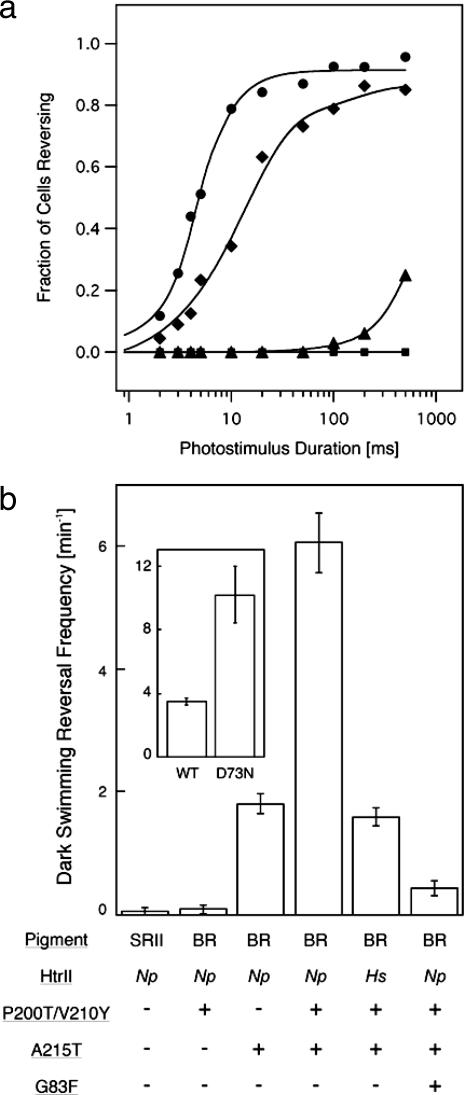
In haloarchaea, light-driven ion transporters have been modified by evolution to produce sensory receptors that relay light signals to transducer proteins controlling motility behavior. The proton pump bacteriorhodopsin and the phototaxis receptor sensory rhodopsin II (SRII) differ by 74% of their residues, with nearly all conserved residues within the photoreactive retinal-binding pocket in the membrane-embedded center of the proteins. Here, we show that three residues in bacteriorhodopsin replaced by the corresponding residues in SRII enable bacteriorhodopsin to efficiently relay the retinal photoisomerization signal to the SRII integral membrane transducer (HtrII) and induce robust phototaxis responses. A single replacement (Ala-215-Thr), bridging the retinal and the membrane-embedded surface, confers weak phototaxis signaling activity, and the additional two (surface substitutions Pro-200-Thr and Val-210-Tyr), expected to align bacteriorhodopsin and HtrII in similar juxtaposition as SRII and HtrII, greatly enhance the signaling. In SRII, the three residues form a chain of hydrogen bonds from the retinal's photoisomerized C(13)=C(14) double bond to residues in the membrane-embedded alpha-helices of HtrII. The results suggest a chemical mechanism for signaling that entails initial storage of energy of photoisomerization in SRII's hydrogen bond between Tyr-174, which is in contact with the retinal, and Thr-204, which borders residues on the SRII surface in contact with HtrII, followed by transfer of this chemical energy to drive structural transitions in the transducer helices. The results demonstrate that evolution accomplished an elegant but simple conversion: The essential differences between transport and signaling proteins in the rhodopsin family are far less than previously imagined.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ruiz-Gonzalez MX, Marin I. J Mol Evol. 2004;58:348–358. - PubMed
-
- Spudich JL, Jung K-H. In: Handbook of Photosensory Receptors. Briggs W, Spudich JL, editors. Weinheim, Germany: Wiley; 2005. pp. 1–24.
-
- Sharma AK, Spudich JL, Doolittle WF. Trends Microbiol. 2006 Sep 26; 10.1016/j.tim.2006.09.006. - PubMed
-
- Luecke H, Lanyi JK. Adv Protein Chem. 2003;63:111–130. - PubMed
-
- Béjà O., Spudich EN, Spudich JL, Leclerc M, DeLong EF. Nature. 2001;411:786–789. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
