

Identified motoneurons involved in sexual and eliminative functions in the rat are powerfully excited by vasopressin and tachykinins
- PMID: 17050711
- PMCID: PMC6674731
- DOI: 10.1523/JNEUROSCI.3364-06.2006
Identified motoneurons involved in sexual and eliminative functions in the rat are powerfully excited by vasopressin and tachykinins
Abstract
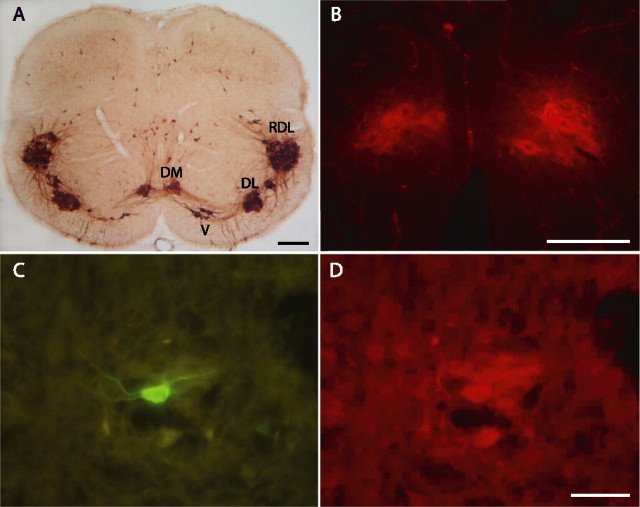
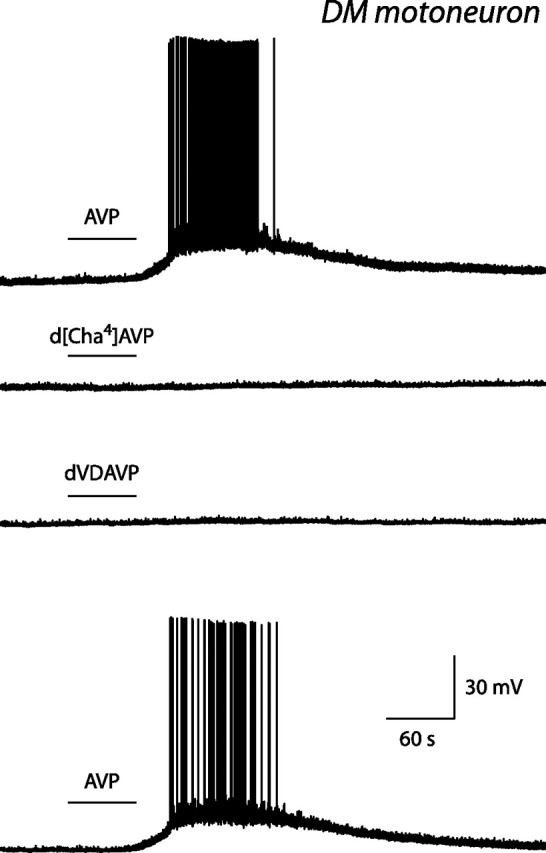
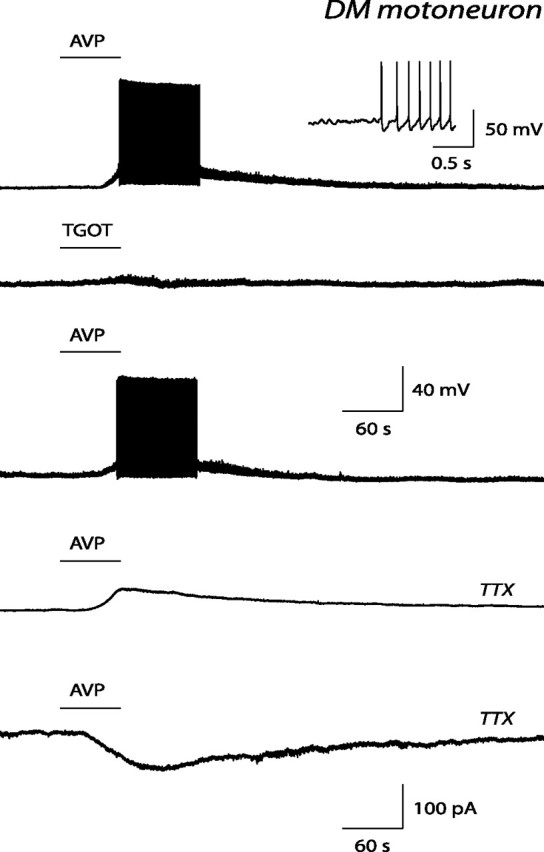
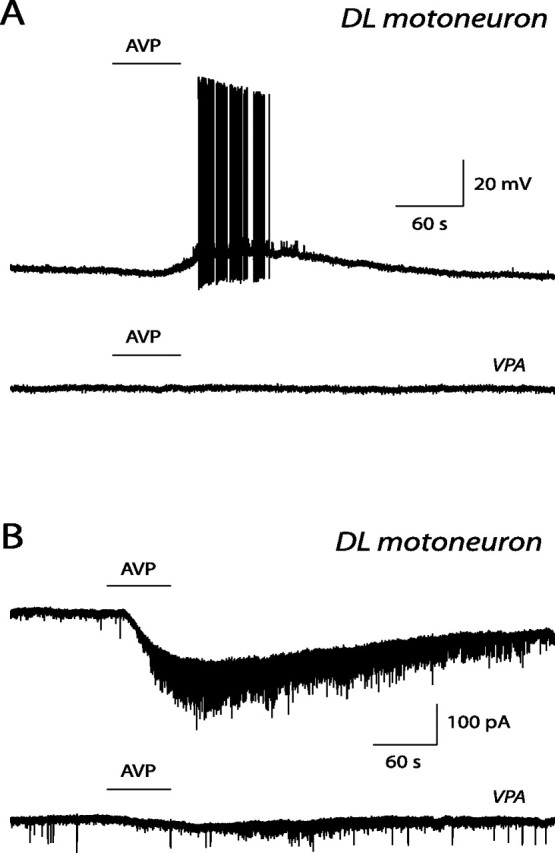
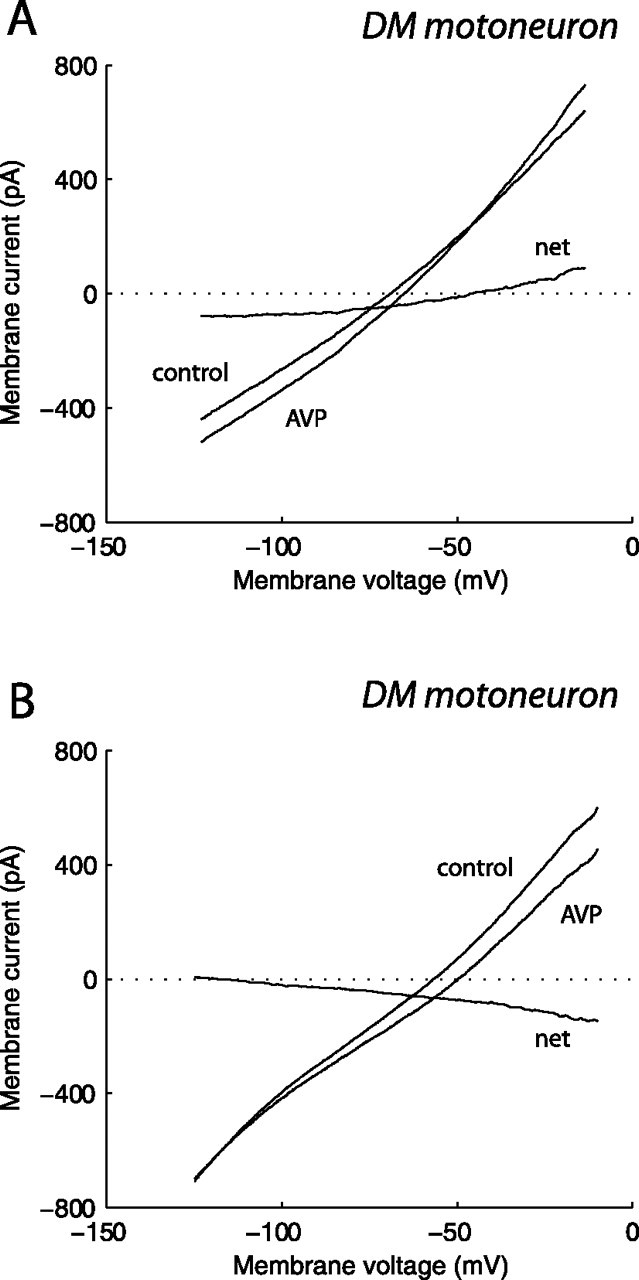
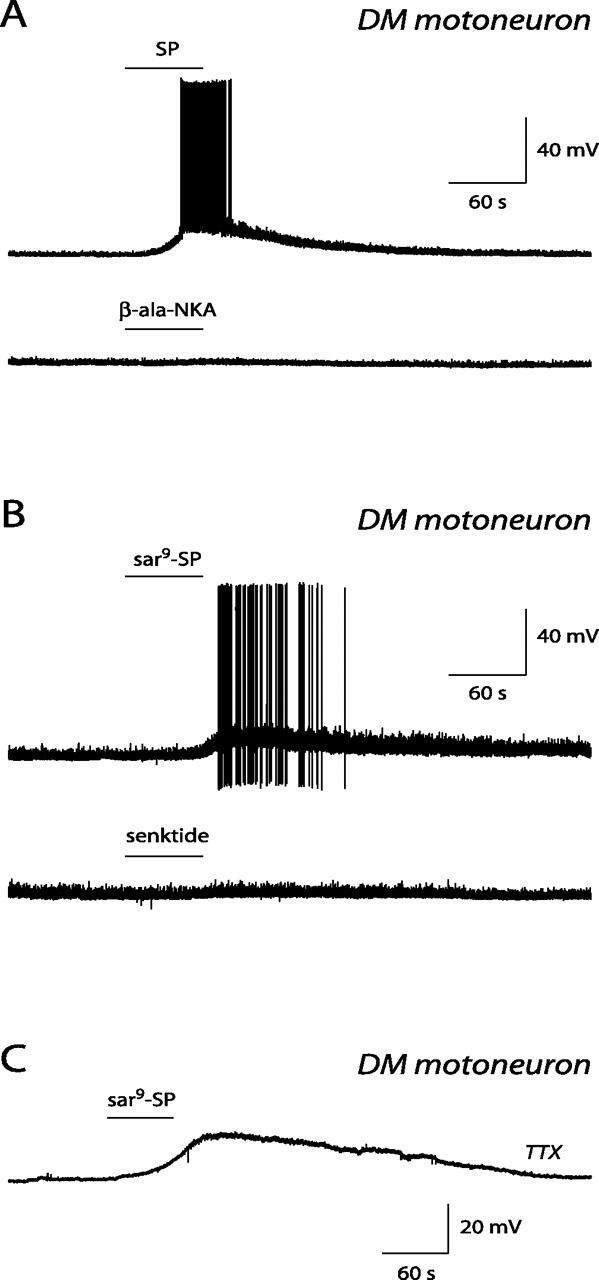
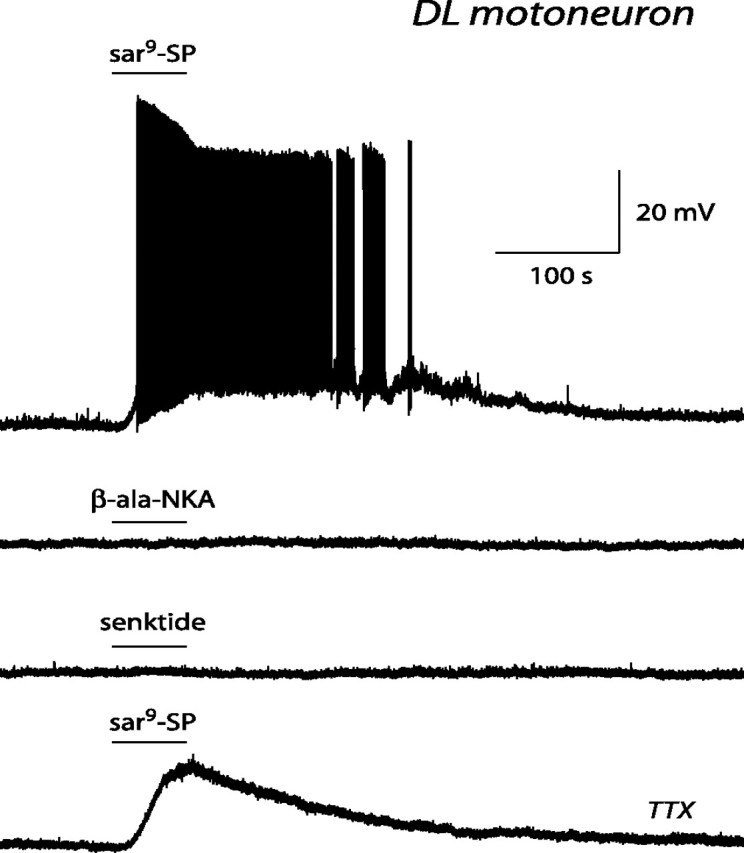
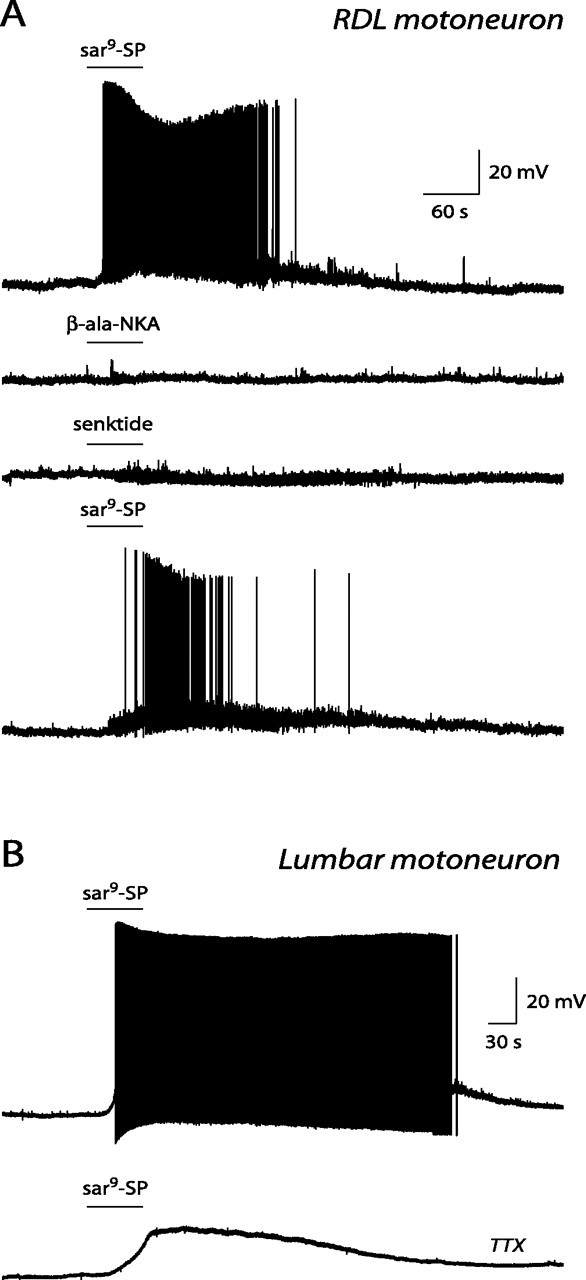
The pudendal motor system is constituted by striated muscles of the pelvic floor and the spinal motoneurons that innervate them. It plays a role in eliminative functions of the bladder and intestine and in sexual function. Pudendal motoneurons are located in the ventral horn of the caudal lumbar spinal cord and send their axon into the pudendal nerve. In the rat, binding sites for vasopressin and tachykinin are present in the dorsomedial and dorsolateral pudendal nuclei, suggesting that these neuropeptides may affect pudendal motoneurons. The aim of the present study was to investigate possible effects of vasopressin and tachykinins on these motoneurons. Recordings were performed in spinal cord slices of young male rats using the whole-cell patch-clamp technique. Before recording, motoneurons were identified by 1,1'-dilinoleyl-3,3,3',3'-tetramethylindocarbocyanine, 4-chlorobenzenesulfonate retrograde labeling. The identification was confirmed, a posteriori, by choline acetyltransferase immunocytochemistry. Vasopressin and tachykinins caused a powerful excitation of pudendal motoneurons. The peptide-evoked depolarization, or the peptide-evoked inward current, persisted in the presence of tetrodotoxin, indicating that these effects were mainly postsynaptic. By using selective receptor agonists and antagonist, we determined that vasopressin acted via vasopressin 1a (V1a), but not V1b, V2, or oxytocin receptors, whereas tachykinins acted via neurokinin 1 (NK1), but not NK2 or NK3, receptors. Vasopressin acted by enhancing a nonselective cationic conductance; in some motoneurons, it also probably suppressed a resting K+ conductance. Our data show that vasopressin and tachykinins can excite pudendal motoneurons and thus influence the force of striated perineal muscles involved in eliminative and sexual functions.
Figures
Similar articles
-
The vasopressin-induced excitation of hypoglossal and facial motoneurons in young rats is mediated by V1a but not V1b receptors, and is independent of intracellular calcium signalling.Eur J Neurosci. 2006 Sep;24(6):1565-74. doi: 10.1111/j.1460-9568.2006.05038.x. Eur J Neurosci. 2006. PMID: 17004920
-
Pudendal motoneurons of the rat located in separated spinal nuclei possess nicotinic acetylcholine receptors having distinct pharmacological profiles.Eur J Neurosci. 2008 Sep;28(5):924-30. doi: 10.1111/j.1460-9568.2008.06403.x. Epub 2008 Aug 8. Eur J Neurosci. 2008. PMID: 18691331
-
Vasopressin facilitates glycinergic and GABAergic synaptic transmission in developing hypoglossal motoneurons.Eur J Neurosci. 2005 Mar;21(6):1601-9. doi: 10.1111/j.1460-9568.2005.03996.x. Eur J Neurosci. 2005. PMID: 15845087
-
Overview of cellular electrophysiological actions of vasopressin.Eur J Pharmacol. 2008 Apr 7;583(2-3):243-54. doi: 10.1016/j.ejphar.2007.11.074. Epub 2008 Jan 30. Eur J Pharmacol. 2008. PMID: 18280467 Review.
-
Vasopressin acting at V1-type receptors produces membrane depolarization in neonatal rat spinal lateral column neurons.Prog Brain Res. 1998;119:275-84. doi: 10.1016/s0079-6123(08)61575-7. Prog Brain Res. 1998. PMID: 10074794 Review.
Cited by
-
Activation of V1a vasopressin receptors excite subicular pyramidal neurons by activating TRPV1 and depressing GIRK channels.Neuropharmacology. 2021 Jun 1;190:108565. doi: 10.1016/j.neuropharm.2021.108565. Epub 2021 Apr 20. Neuropharmacology. 2021. PMID: 33891950 Free PMC article.
-
Neural control of the lower urinary tract: peripheral and spinal mechanisms.Neurourol Urodyn. 2010;29(1):128-39. doi: 10.1002/nau.20837. Neurourol Urodyn. 2010. PMID: 20025024 Free PMC article. Review.
-
Expression of arginine vasotocin receptors in the developing zebrafish CNS.Gene Expr Patterns. 2013 Dec;13(8):335-42. doi: 10.1016/j.gep.2013.06.005. Epub 2013 Jul 2. Gene Expr Patterns. 2013. PMID: 23830982 Free PMC article.
-
Voluntary urination control by brainstem neurons that relax the urethral sphincter.Nat Neurosci. 2018 Sep;21(9):1229-1238. doi: 10.1038/s41593-018-0204-3. Epub 2018 Aug 13. Nat Neurosci. 2018. PMID: 30104734 Free PMC article.
-
Neural control of the female urethral and anal rhabdosphincters and pelvic floor muscles.Am J Physiol Regul Integr Comp Physiol. 2010 Aug;299(2):R416-38. doi: 10.1152/ajpregu.00111.2010. Epub 2010 May 19. Am J Physiol Regul Integr Comp Physiol. 2010. PMID: 20484700 Free PMC article. Review.
References
-
- Aghajanian GK, Rasmussen K. Intracellular studies in the facial nucleus illustrating a simple new method for obtaining viable motoneurons in adult rat brain slices. Synapse. 1989;3:331–338. - PubMed
-
- Alberi S, Dubois-Dauphin M, Dreifuss JJ, Raggenbass M. Modulation by divalent cations of the current generated by vasopressin in facial motoneurons. Brain Res. 1993;624:326–330. - PubMed
-
- Beresford IJ, Ireland SJ, Stables J, Hagan RM. Ontogeny and characterization of [125I]Bolton Hunter-eledoisin binding sites in rat spinal cord by quantitative autoradiography. Neuroscience. 1992;46:225–232. - PubMed
-
- Breedlove SM, Arnold AP. Hormone accumulation in a sexually dimorphic motor nucleus of the rat spinal cord. Science. 1980;210:564–566. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources