

Chondroitinase ABC promotes sprouting of intact and injured spinal systems after spinal cord injury
- PMID: 17050723
- PMCID: PMC3339436
- DOI: 10.1523/JNEUROSCI.2980-06.2006
Chondroitinase ABC promotes sprouting of intact and injured spinal systems after spinal cord injury
Abstract
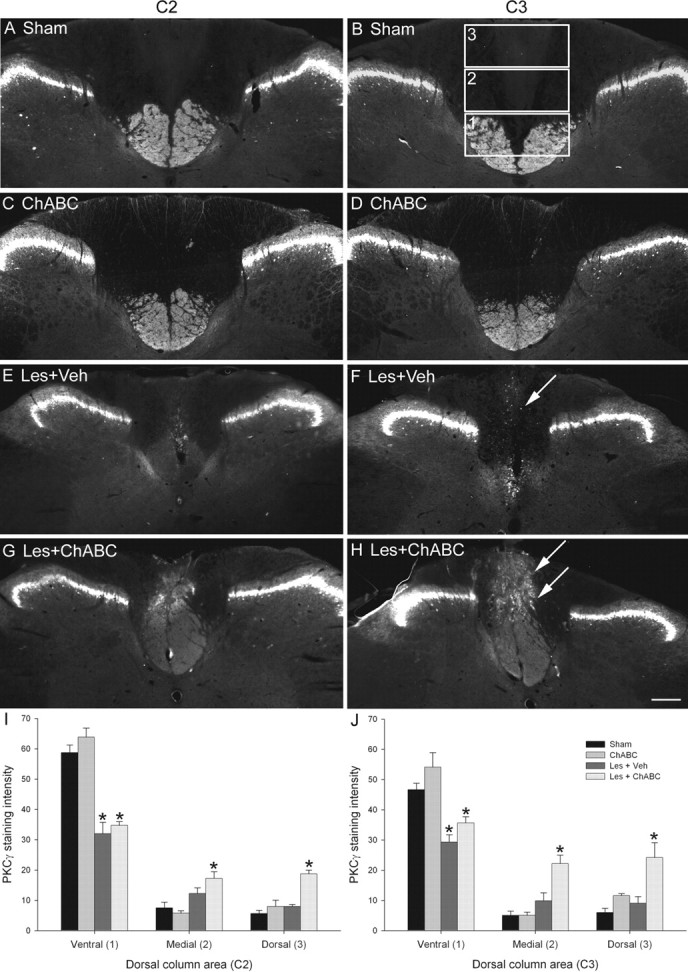
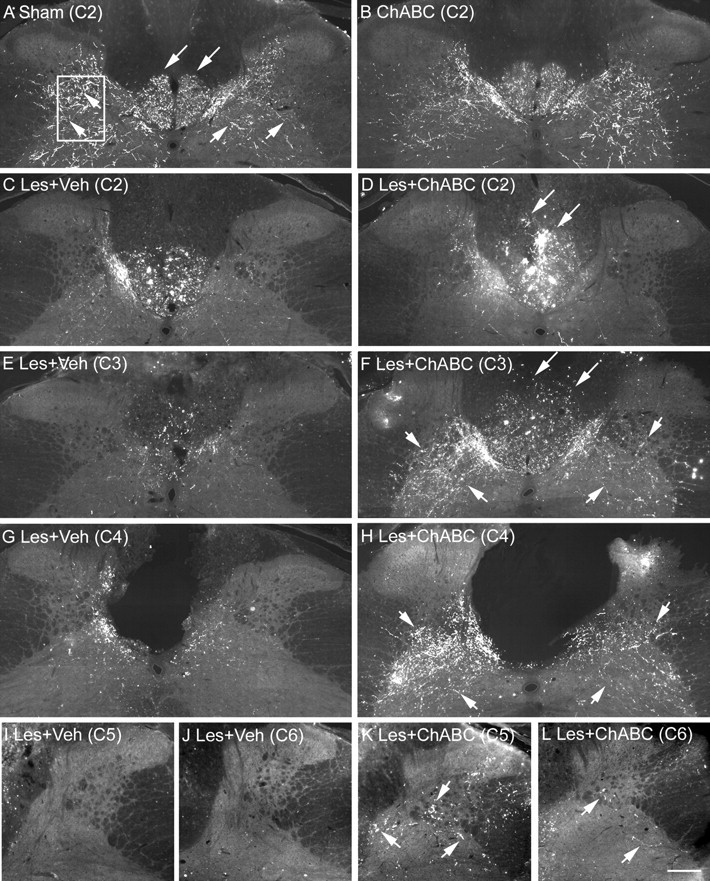
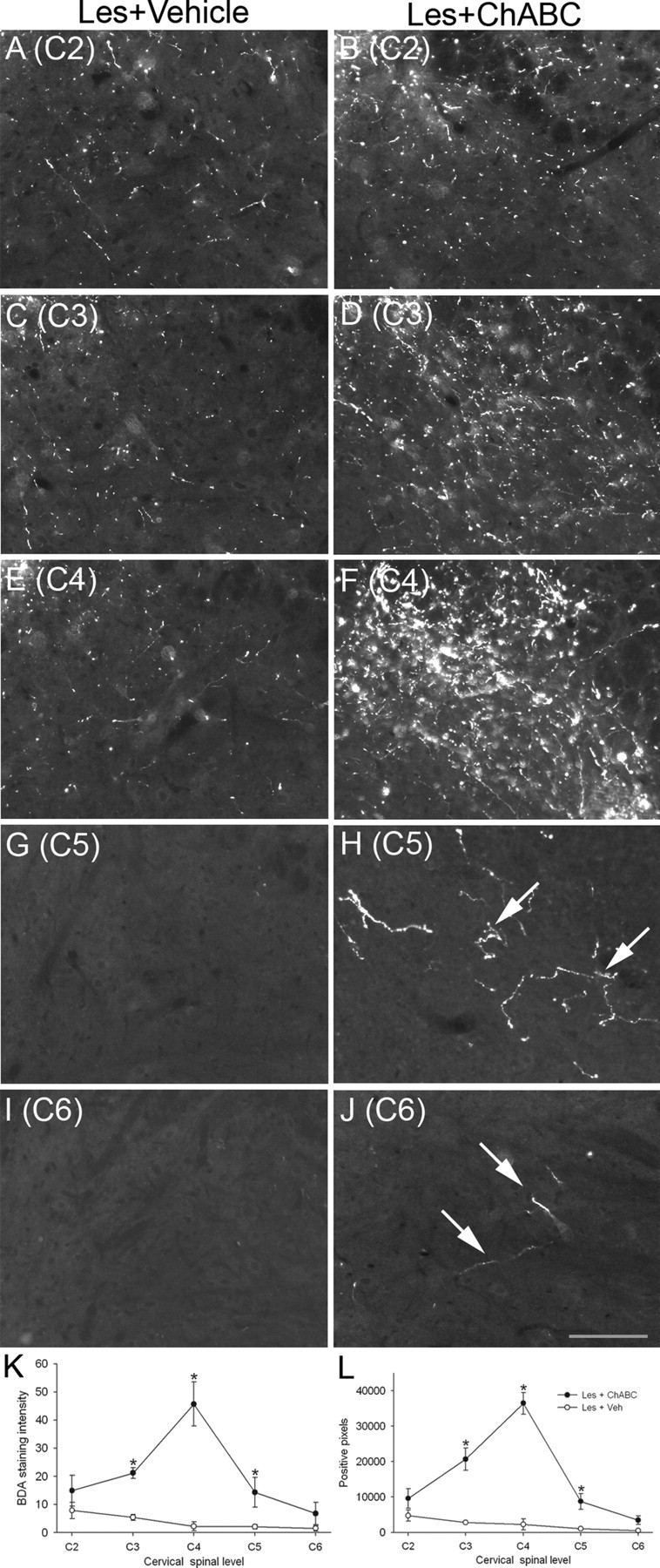
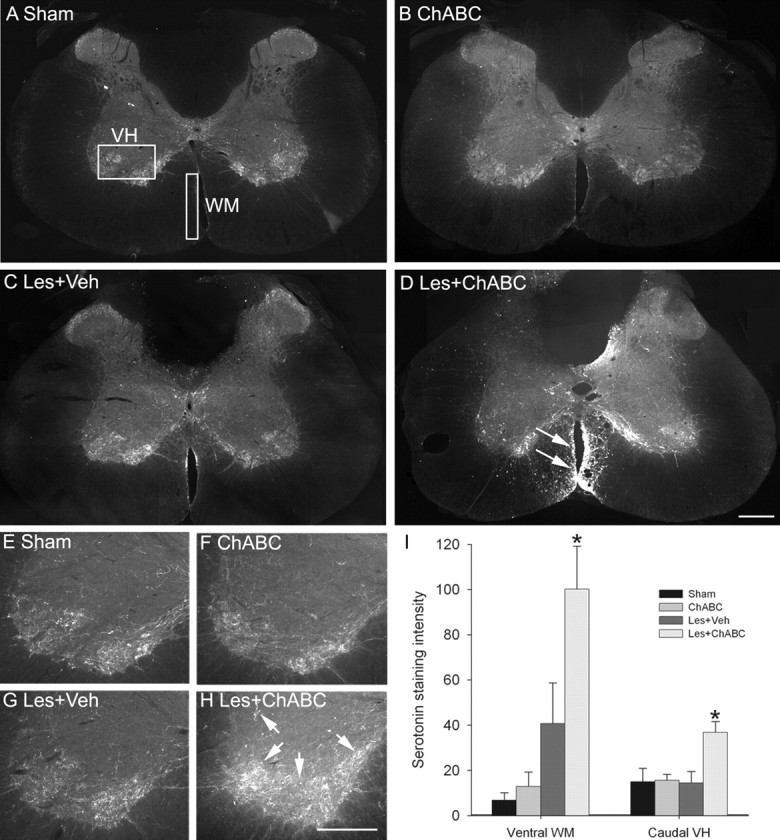
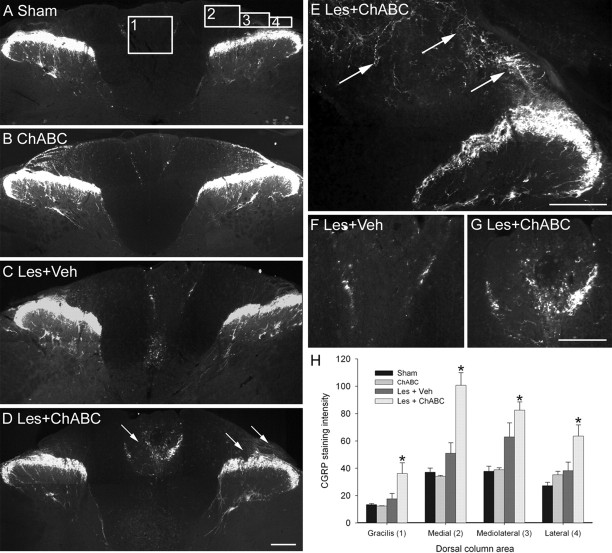
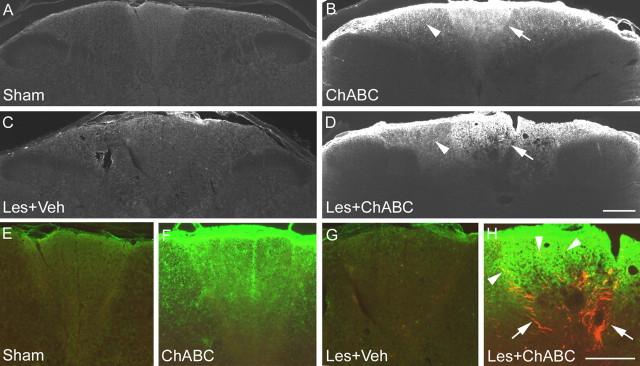
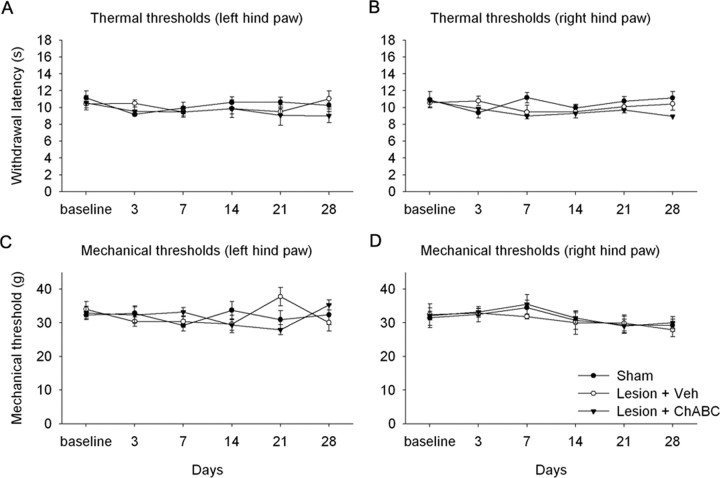
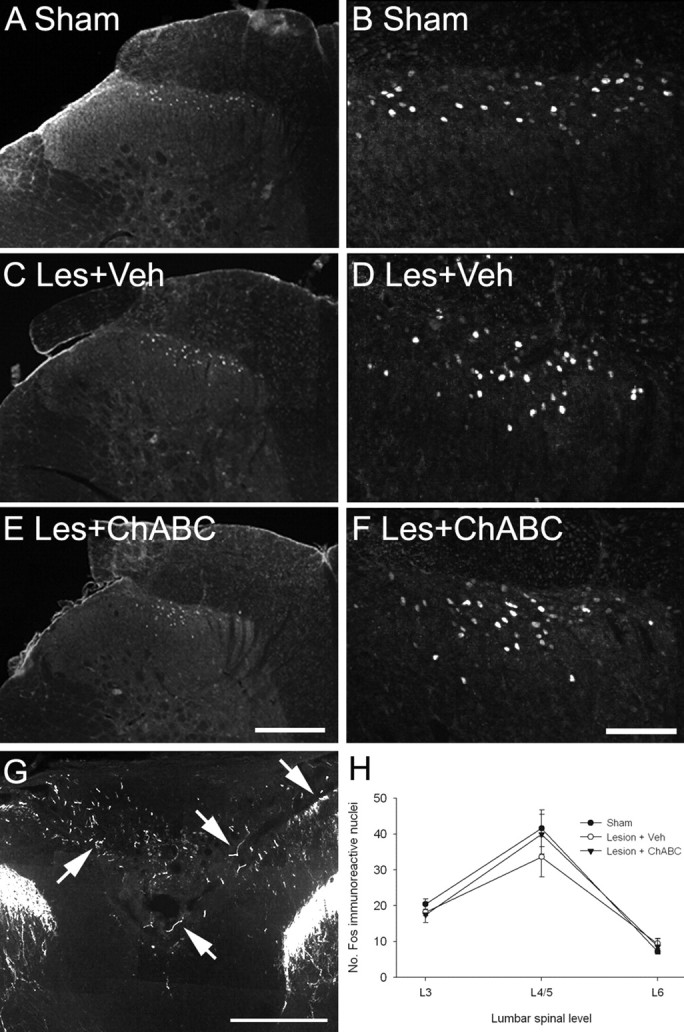
Chondroitin sulfate proteoglycans (CSPGs) are inhibitory extracellular matrix molecules that are upregulated after CNS injury. Degradation of CSPGs using the enzyme chondroitinase ABC (ChABC) can promote functional recovery after spinal cord injury. However, the mechanisms underlying this recovery are not clear. Here we investigated the effects of ChABC treatment on promoting plasticity within the spinal cord. We found robust sprouting of both injured (corticospinal) and intact (serotonergic) descending projections as well as uninjured primary afferents after a cervical dorsal column injury and ChABC treatment. Sprouting fibers were observed in aberrant locations in degenerating white matter proximal to the injury in regions where CSPGs had been degraded. Corticospinal and serotonergic sprouting fibers were also observed in spinal gray matter at and below the level of the lesion, indicating increased innervation in the terminal regions of descending projections important for locomotion. Spinal-injured animals treated with a vehicle solution showed no significant sprouting. Interestingly, ChABC treatment in uninjured animals did not induce sprouting in any system. Thus, both denervation and CSPG degradation were required to promote sprouting within the spinal cord. We also examined potential detrimental effects of ChABC-induced plasticity. However, although primary afferent sprouting was observed after lumbar dorsal column lesions and ChABC treatment, there was no increased connectivity of nociceptive neurons or development of mechanical allodynia or thermal hyperalgesia. Thus, CSPG digestion promotes robust sprouting of spinal projections in degenerating and denervated areas of the spinal cord; compensatory sprouting of descending systems could be a key mechanism underlying functional recovery.
Figures
References
-
- Bareyre FM, Kerschensteiner M, Raineteau O, Mettenleiter TC, Weinmann O, Schwab ME. The injured spinal cord spontaneously forms a new intraspinal circuit in adult rats. Nat Neurosci. 2004;7:269–277. - PubMed
-
- Bennett AD, Chastain KM, Hulsebosch CE. Alleviation of mechanical and thermal allodynia by CGRP(8-37) in a rodent model of chronic central pain. Pain. 2000;86:163–175. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical