

Adenosine inhibits cytosolic calcium signals and chemotaxis in hepatic stellate cells
- PMID: 17053161
- PMCID: PMC3224076
- DOI: 10.1152/ajpgi.00208.2006
Adenosine inhibits cytosolic calcium signals and chemotaxis in hepatic stellate cells
Abstract
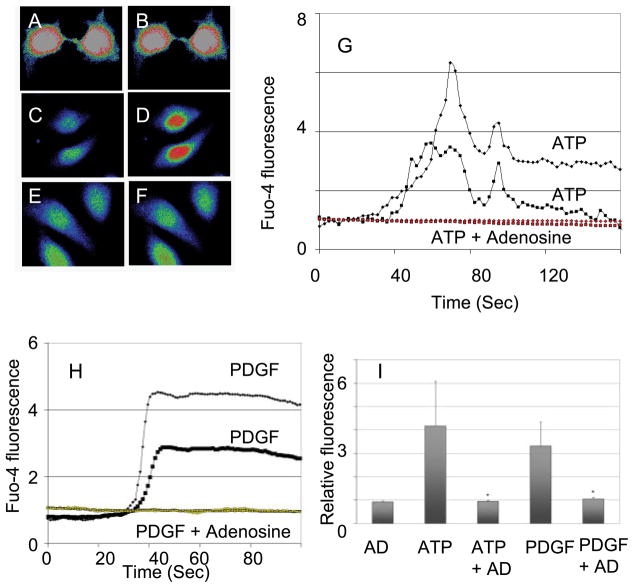
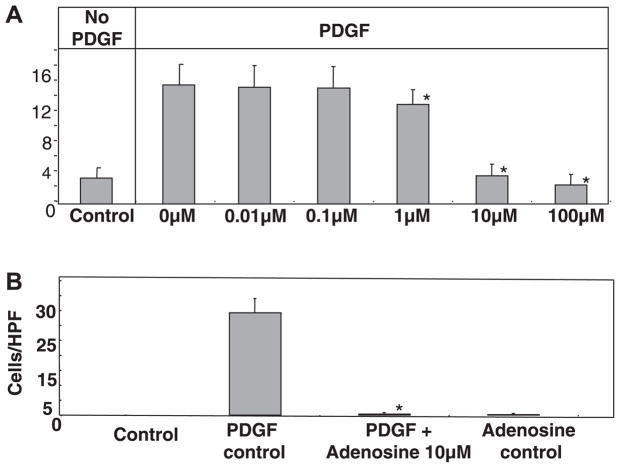
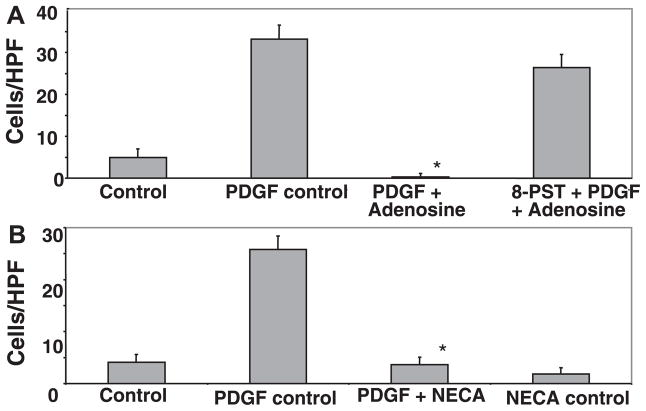
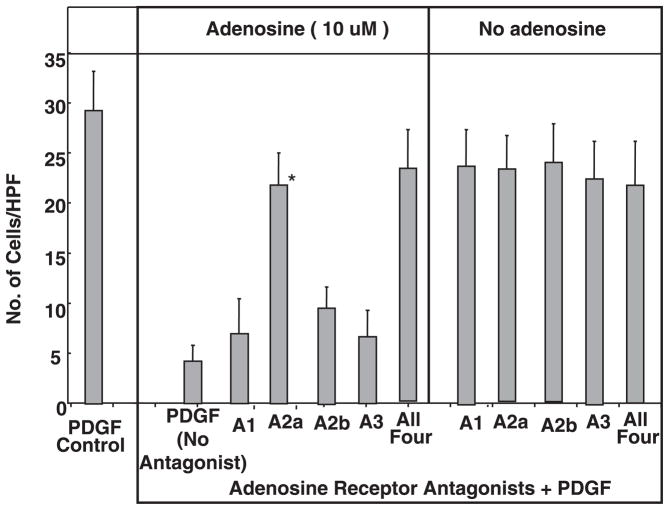
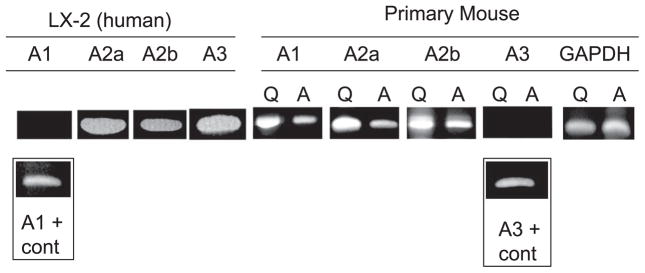
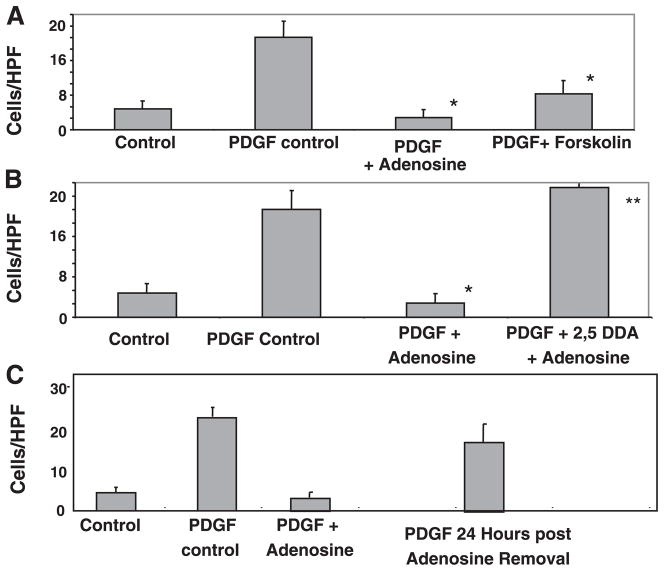
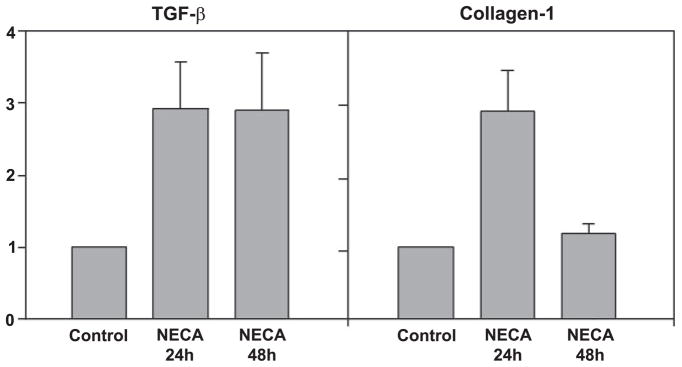
Adenosine is produced during cellular hypoxia and apoptosis, resulting in elevated tissue levels at sites of injury. Adenosine is also known to regulate a number of cellular responses to injury, but its role in hepatic stellate cell (HSC) biology and liver fibrosis is poorly understood. We tested the effect of adenosine on the cytosolic Ca2+ concentration, chemotaxis, and upregulation of activation markers in HSCs. We showed that adenosine did not induce an increase in the cytosolic Ca2+ concentration in LX-2 cells and, in addition, inhibited increases in the cytosolic Ca2+ concentration in response to ATP and PDGF. Using a Transwell system, we showed that adenosine strongly inhibited PDGF-induced HSC chemotaxis in a dose-dependent manner. This inhibition was mediated via the A(2a) receptor, was reversible, was reproduced by forskolin, and was blocked by the adenylate cyclase inhibitor 2,5-dideoxyadenosine. Adenosine also upregulated the production of TGF-beta and collagen I mRNA. In conclusion, adenosine reversibly inhibits Ca2+ fluxes and chemotaxis of HSCs and upregulates TGF-beta and collagen I mRNA. We propose that adenosine provides 1) a "stop" signal to HSCs when they reach sites of tissue injury with high adenosine concentrations and 2) stimulates transdifferentiation of HSCs by upregulating collagen and TGF-beta production.
Figures
References
-
- Aguirre-Ghiso JA, Estrada Y, Liu D, Ossowski L. ERK (MAPK) activity as a determinant of tumor growth and dormancy; regulation by p38 (SAPK) Cancer Res. 2003;63:1684–1695. - PubMed
-
- Albanis E, Friedman SL. Hepatic fibrosis. Pathogenesis and principles of therapy. Clin Liver Dis. 2001;5:315–334. - PubMed
-
- Bonner JC. Regulation of PDGF and its receptors in fibrotic diseases. Cytokine Growth Factor Rev. 2004;15:255–273. - PubMed
-
- Dawson AP. Calcium signalling: how do IP3 receptors work? Curr Biol. 1997;7:R544–R547. - PubMed
-
- Friedman SL. Mechanisms of disease: mechanisms of hepatic fibrosis and therapeutic implications. Nat Clin Pract Gastroenterol Hepatol. 2004;1:98–105. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous