

Nucleocytoplasmic shuttling of the Rpb4p and Rpb7p subunits of Saccharomyces cerevisiae RNA polymerase II by two pathways
- PMID: 17056745
- PMCID: PMC1694818
- DOI: 10.1128/EC.00288-06
Nucleocytoplasmic shuttling of the Rpb4p and Rpb7p subunits of Saccharomyces cerevisiae RNA polymerase II by two pathways
Abstract
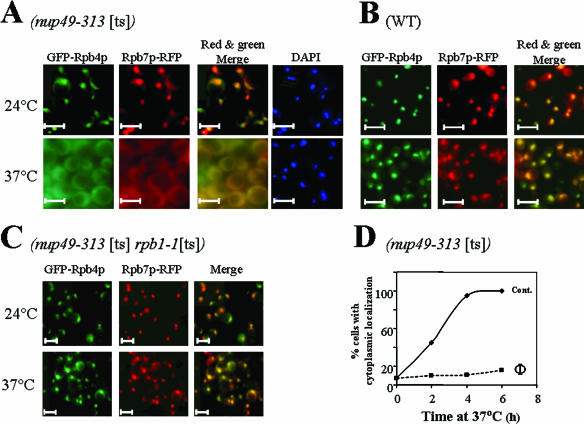
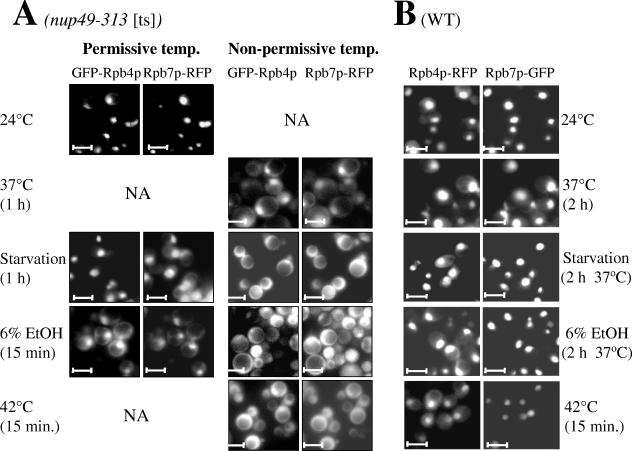
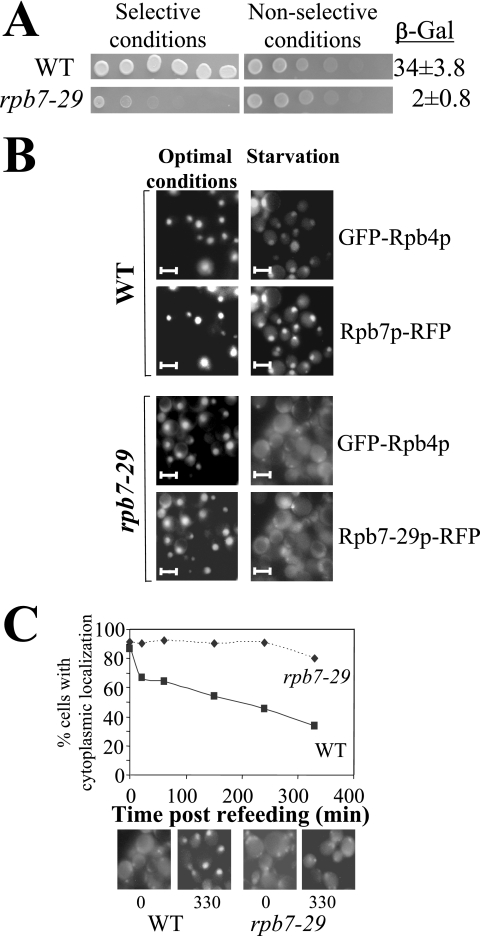
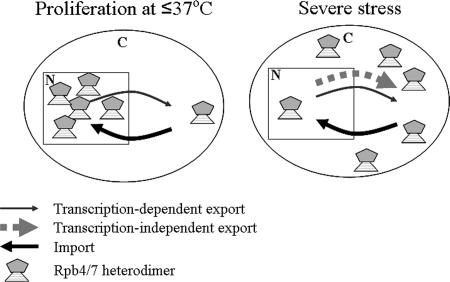
Rpb4p and Rpb7p are subunits of the RNA polymerase II of Saccharomyces cerevisiae that form a dissociable heterodimeric complex. Whereas the only reported function of Rpb7p is related to transcription, Rpb4p has been found to also act in mRNA export and in the major mRNA decay pathway that operates in the cytoplasm, thus raising the possibility that Rpb4p links between the nuclear and cytoplasmic processes. Here we show that both Rpb4p and Rpb7p shuttle between the nucleus and the cytoplasm. Shuttling kinetics of the two proteins are similar as long as their interaction is possible, suggesting that they shuttle as a heterodimer. Under normal conditions, shuttling of Rpb4p and Rpb7p depends on ongoing transcription. However, during severe stresses of heat shock, ethanol, and starvation, the two proteins shuttle via a transcription-independent pathway. Thus, Rpb4p and Rpb7p shuttle via two pathways, depending on environmental conditions.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
