

Two unlinked double-strand breaks can induce reciprocal exchanges in plant genomes via homologous recombination and nonhomologous end joining
- PMID: 17057227
- PMCID: PMC1775016
- DOI: 10.1534/genetics.106.065185
Two unlinked double-strand breaks can induce reciprocal exchanges in plant genomes via homologous recombination and nonhomologous end joining
Abstract
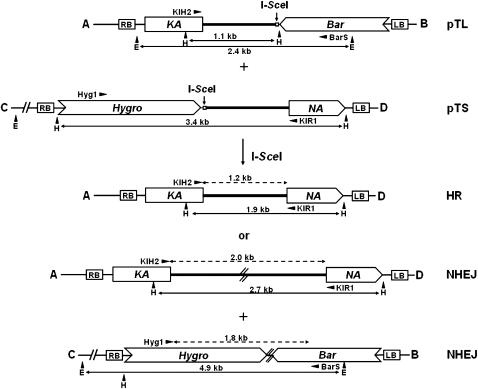
Using the rare-cutting endonuclease I-SceI we were able to demonstrate before that the repair of a single double-strand break (DSB) in a plant genome can be mutagenic due to insertions and deletions. However, during replication or due to irradiation several breaks might be induced simultaneously. To analyze the mutagenic potential of such a situation we established an experimental system in tobacco harboring two unlinked transgenes, each carrying an I-SceI site. After transient expression of I-SceI a kanamycin-resistance marker could be restored by joining two previously unlinked broken ends, either by homologous recombination (HR) or by nonhomologous end joining (NHEJ). Indeed, we were able to recover HR and NHEJ events with similar frequencies. Despite the fact that no selection was applied for joining the two other ends, the respective linkage could be detected in most cases tested, demonstrating that the respective exchanges were reciprocal. The frequencies obtained indicate that DSB-induced translocation is up to two orders of magnitude more frequent in somatic cells than ectopic gene conversion. Thus, DSB-induced reciprocal exchanges might play a significant role in plant genome evolution. The technique applied in this study may also be useful for the controlled exchange of unlinked sequences in plant genomes.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
