

Multiple steps determine the overall rate of the reduction of 5alpha-dihydrotestosterone catalyzed by human type 3 3alpha-hydroxysteroid dehydrogenase: implications for the elimination of androgens
- PMID: 17059222
- PMCID: PMC2597410
- DOI: 10.1021/bi060591r
Multiple steps determine the overall rate of the reduction of 5alpha-dihydrotestosterone catalyzed by human type 3 3alpha-hydroxysteroid dehydrogenase: implications for the elimination of androgens
Abstract
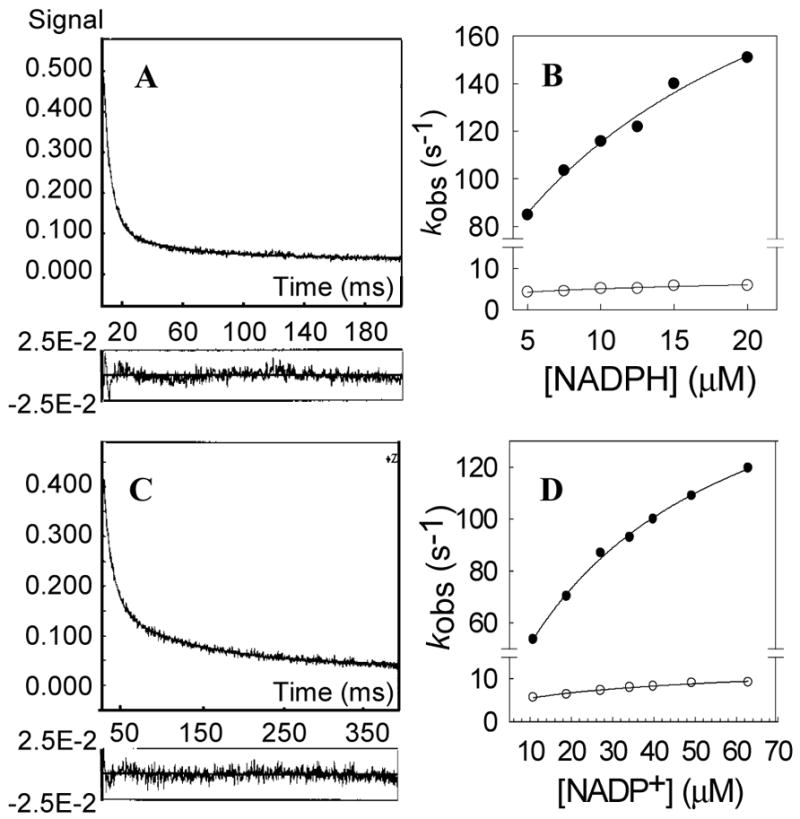
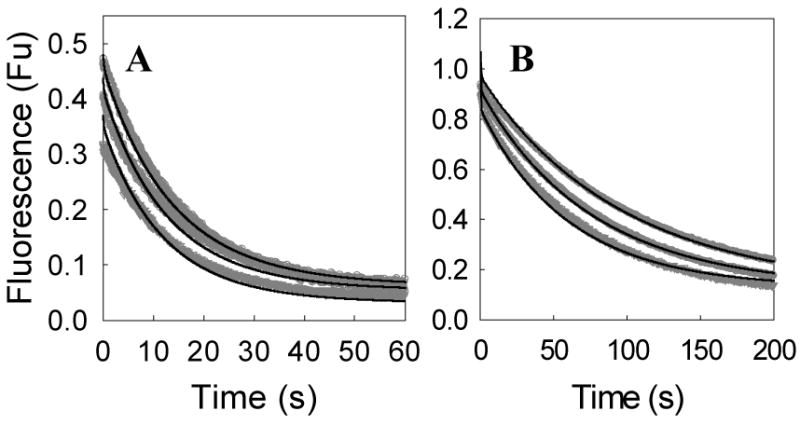
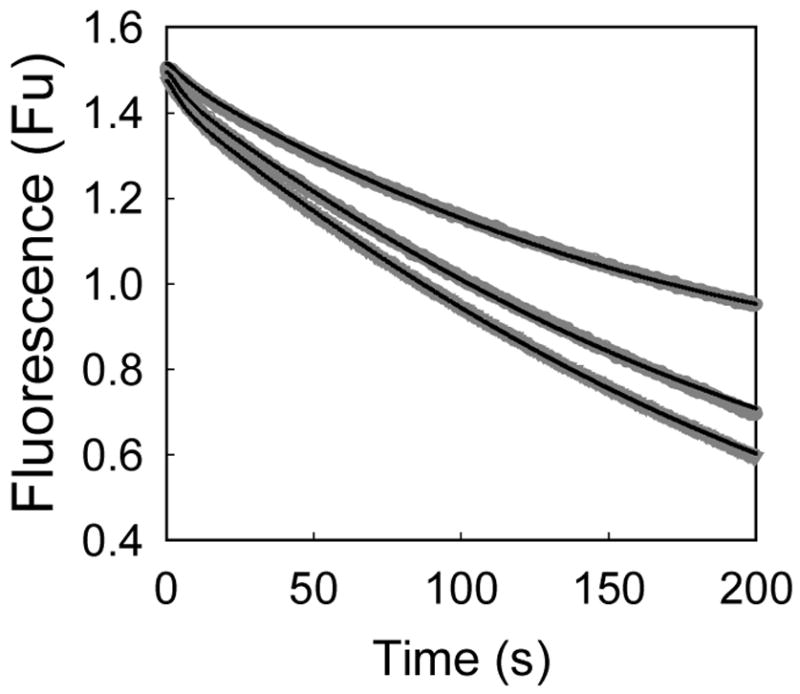
Human type 3 3alpha-hydroxysteroid dehydrogenase, or aldo-keto reductase (AKR) 1C2, eliminates the androgen signal in human prostate by reducing 5alpha-dihydrotestosterone (DHT, potent androgen) to form 3alpha-androstanediol (inactive androgen), thereby depriving the androgen receptor of its ligand. The k(cat) for the NADPH-dependent reduction of DHT catalyzed by AKR1C2 is 0.033 s(-1). We employed transient kinetics and kinetic isotope effects to dissect the contribution of discrete steps to this low k(cat) value. Stopped-flow experiments to measure the formation of the AKR1C2.NADP(H) binary complex indicated that two slow isomerization events occur to yield a tight complex. A small primary deuterium isotope effect on k(cat) (1.5) and a slightly larger effect on k(cat)/K(m) (2.1) were observed in the steady state. In the transient state, the maximum rate constant for the single turnover of DHT (k(trans)) was determined to be 0.11 s(-1) for the NADPH-dependent reaction, which was approximately 4-fold greater than the corresponding k(cat) x k(trans) was significantly reduced when NADPD was substituted for NADPH, resulting in an apparent (D)k(trans) of 3.5. Thus, the effects of isotopic substitution on the hydride transfer step were masked by slow events that follow or precede the chemical transformation. Transient multiple-turnover reactions generated curvilinear reaction traces, consistent with the product formation and release occurring at comparable rates. Global fitting analysis of the transient kinetic data enabled the estimate of the rate constants for the three-step cofactor binding/release model and for the minimal ordered bi-bi turnover mechanism. Results were consistent with a kinetic mechanism in which a series of slow events, including the chemical step (0.12 s(-1)), the release of the steroid product (0.081 s(-1)), and the release of the cofactor product (0.21 s(-1)), combine to yield the overall observed low turnover number.
Figures





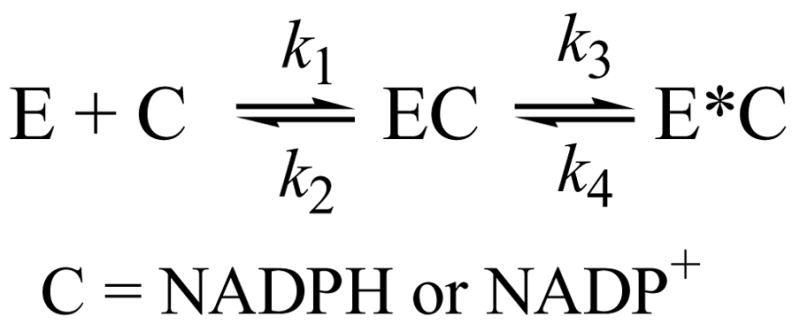
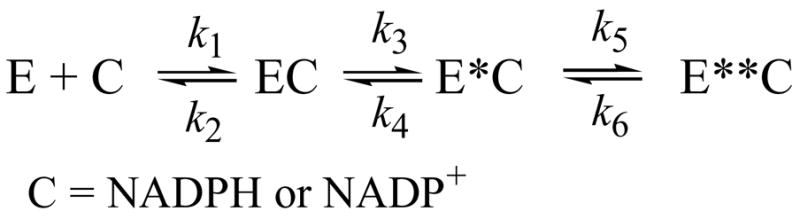

References
-
- Penning TM, Jin Y, Steckelbroeck S, Rizner TL, Lewis M. Structure-function of human 3α-hydroxysteroid dehydrogenases: genes and proteins. Mol Cell Endocrinol. 2004;215:63–72. - PubMed
-
- Jez JM, Flynn TG, Penning TM. A new nomenclature for the aldo-keto reductase superfamily. Biochem Pharmacol. 1997;54:639–647. - PubMed
-
- Hyndman D, Bauman DR, Heredia VV, Penning TM. The aldo-keto reductase superfamily homepage. Chem Biol Interact. 2003;143-144:621–631. - PubMed
-
- Penning TM. Molecular endocrinology of hydroxysteroid dehydrogenases. Endocrine Reviews. 1997;18:281–305. - PubMed
-
- Penning TM. Hydroxysteroid dehydrogenases and pre-receptor regulation of steroid hormone action. Hum Reprod Update. 2003;9:193–205. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
