

Role of monocyte-acquired hemozoin in suppression of macrophage migration inhibitory factor in children with severe malarial anemia
- PMID: 17060471
- PMCID: PMC1828375
- DOI: 10.1128/IAI.01327-06
Role of monocyte-acquired hemozoin in suppression of macrophage migration inhibitory factor in children with severe malarial anemia
Abstract
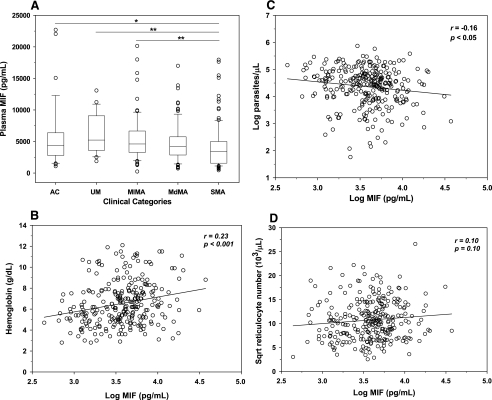
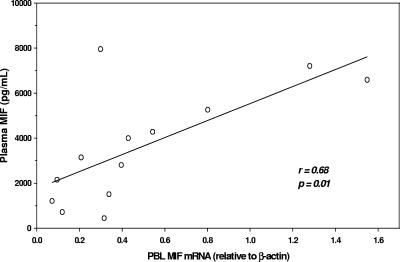
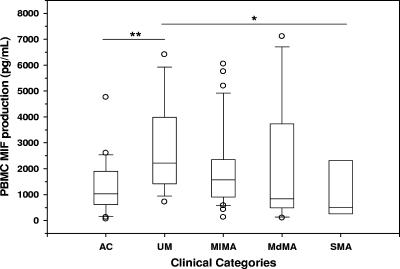
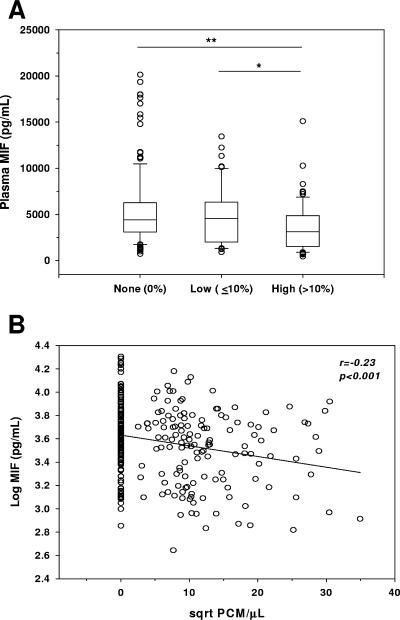
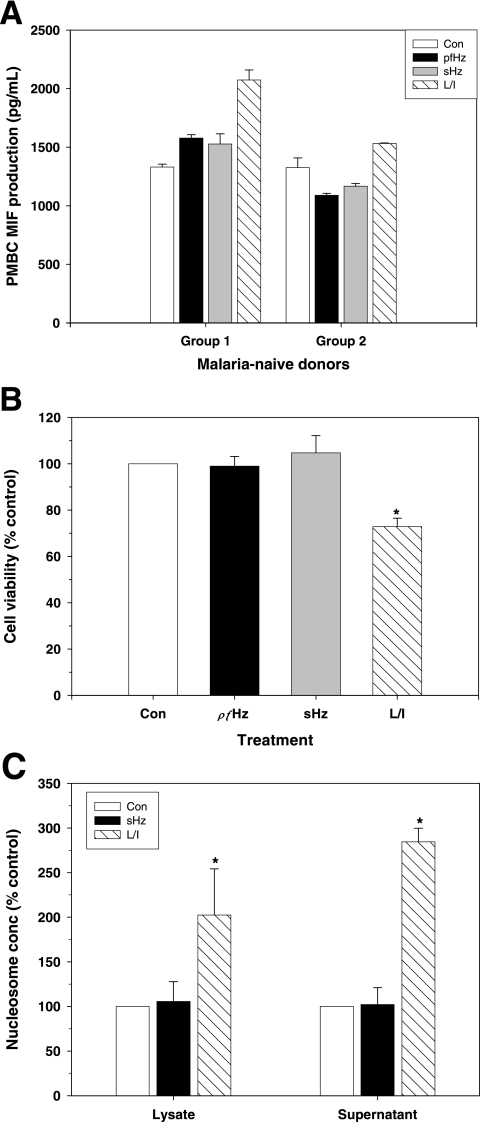
Severe malarial anemia (SMA), caused by Plasmodium falciparum infections, is one of the leading causes of childhood mortality in sub-Saharan Africa. Although the molecular determinants of SMA are largely undefined, dysregulation in host-derived inflammatory mediators influences disease severity. Macrophage migration inhibitory factor (MIF) is an important regulator of innate inflammatory responses that has recently been shown to suppress erythropoiesis and promote pathogenesis of SMA in murine models. To examine the role of MIF in the development of childhood SMA, peripheral blood MIF production was examined in Kenyan children (aged <3 years, n = 357) with P. falciparum malarial anemia. All children in the study were free from bacteremia and human immunodeficiency virus type 1. Since deposition of malarial pigment (hemozoin [Hz]) contributes to suppression of erythropoiesis, the relationship between MIF concentrations and monocytic acquisition of Hz was also examined in vivo and in vitro. Circulating MIF concentrations declined with increasing severity of anemia and significantly correlated with peripheral blood leukocyte MIF transcripts. However, MIF concentrations in peripheral blood were not significantly associated with reticulocyte production. Multivariate regression analyses, controlling for age, gender, and parasitemia, further revealed that elevated levels of pigment-containing monocytes (PCM) was associated with SMA and decreased MIF production. In addition, PCM levels were a better predictor of hemoglobin and MIF concentrations than parasite density. Additional experiments in malaria-naive individuals demonstrated that hemozoin caused both increased and decreased MIF production in cultured peripheral blood mononuclear cells (PBMC) in a donor-specific manner, independent of apoptosis. However, PBMC MIF production in children with acute malaria progressively declined with increasing anemia severity. Results presented here demonstrate that acquisition of hemozoin by monocytes is associated with suppression of peripheral blood MIF production and enhanced severity of anemia in childhood malaria.
Figures
References
-
- Abdalla, S. H. 1990. Hematopoiesis in human malaria. Blood Cells 16:401-419. - PubMed
-
- Akpede, G. O., and R. M. Sykes. 1993. Malaria with bacteraemia in acutely febrile preschool children without localizing signs: coincidence or association/complication? J. Trop. Med. Hyg. 96:146-150. - PubMed
-
- Alourfi, Z., R. P. Donn, A. Stevens, A. Berry, A. McMaster, and D. W. Ray. 2005. Glucocorticoids suppress macrophage migration inhibitory factor (MIF) expression in a cell-type-specific manner. J. Mol. Endocrinol. 34:583-595. - PubMed
-
- Amodu, O. K., A. A. Adeyemo, P. E. Olumese, and R. A. Gbadegesin. 1998. Intraleucocytic malaria pigment and clinical severity of malaria in children. Trans. R. Soc. Trop. Med. Hyg. 92:54-56. - PubMed
-
- Arese, P., and E. Schwarzer. 1997. Malarial pigment (haemozoin): a very active ‘inert’ substance. Ann. Trop. Med. Parasitol. 91:501-516. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
