

doi: 10.1083/jcb.200605037.
MyoD-positive epiblast cells regulate skeletal muscle differentiation in the embryo
Affiliations
- PMID: 17060497
- PMCID: PMC2064569
- DOI: 10.1083/jcb.200605037
Item in Clipboard
MyoD-positive epiblast cells regulate skeletal muscle differentiation in the embryo
J Cell Biol.
.
Abstract
MyoD mRNA is expressed in a subpopulation of cells within the embryonic epiblast. Most of these cells are incorporated into somites and synthesize Noggin. Ablation of MyoD-positive cells in the epiblast subsequently results in the herniation of organs through the ventral body wall, a decrease in the expression of Noggin, MyoD, Myf5, and myosin in the somites and limbs, and an increase in Pax-3-positive myogenic precursors. The addition of Noggin lateral to the somites compensates for the loss of MyoD-positive epiblast cells. Skeletal muscle stem cells that arise in the epiblast are utilized in the somites to promote muscle differentiation by serving as a source of Noggin.
Figures
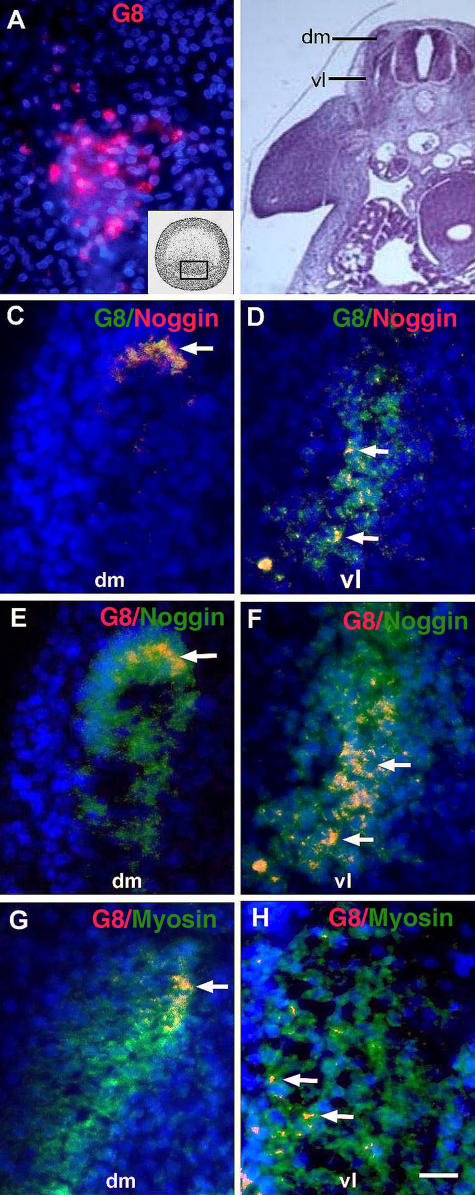
Expression of Noggin and myosin in MyoDpos cells originating in the epiblast. MyoDpos cells labeled with the G8 mAb were present in the posterior region of the stage 2 epiblast (red cells in A). 4–5 d after labeling with G8, stage 25 embryos were examined for expression of Noggin and sarcomeric myosin. Regions indicated on the right side of the embryo in the hematoxylin and eosin–stained section are shown at higher magnification in fluorescence photomicrographs of merged images of G8 mAb (labeled with Alexa Fluor 488 [green in C and D] and rhodamine [red in E–H]) and either Cy3/red-labeled dendrimers to Noggin mRNA (C and D) or Alexa Fluor 488/green–labeled antibodies to Noggin (E and F) or myosin (G and H). Nuclei were stained with Hoechst dye (blue). Double-labeled cells (overlay of red and green) appear yellow (arrows). (C–F) G8pos/ Nogginpos cells were observed in the dorsomedial (dm) and ventrolateral (vl) dermomyotome and myotome. (G and H) G8pos/myosinpos cells were present in the myotome. Bar: (A and C–H) 9 μm; (B) 135 μm.
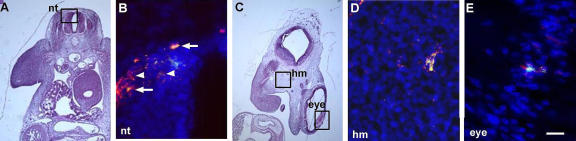
Expression of Noggin in G8pos epiblast cells incorporated into the head and neural tube. (A and C) Areas outlined in hematoxylin and eosin–stained sections are shown at higher magnification in fluorescence photomicrographs that are the merged images of Hoechst-stained nuclei, Alexa Fluor 488–labeled G8 mAb, and Cy3-labeled dendrimers to Noggin mRNA. Double-labeled cells (overlay of red and green) appear yellow. G8pos cells expressed Noggin mRNA in the neural tube (nt; B, arrows), head mesenchyme (hm; D), and eye (E) of the stage 25 embryo. G8neg/Nogginpos cells were present in the neural tube (arrowheads). Bar: (A and C) 135 μm; (B, D, and E) 9 μm.

Morphological effects of ablating MyoDpos/G8pos cells in the epiblast. Stage 2 embryos were treated with the G8 or E12 mAb and complement (G8/cp and E12/cp), or complement (cp) or Hanks' buffer (bc) only. Embryos were incubated in trypan blue to reveal dead cells (blue cells and arrows in A–D). Other embryos were grown for 5 (E–G) or 7 d (H; stages 26 and 30, respectively). Sections were stained with hematoxylin and eosin (I–L). The ventral body wall was discontinuous (E, H, and I, arrows) and the dermatome (J–L, arrows) and dermomyotome/myotome (dm; J, double-headed arrow; K and L, diamond) were thickened in G8/complement-treated embryos (J) compared with embryos treated with buffer alone (K) or E12 and complement (L). G8/complement-treated embryos also exhibited malformations of the eye (F and H, black arrowheads) and facial prominences (E, white arrowhead). The notochord (nc) and neural tube (nt) appeared normal in G8/complement-treated embryos (J). The neural tube was kinked in E12/complement-treated embryos (L). Bar: (A–D and J–L) 56 μm; (I) 135 μm.
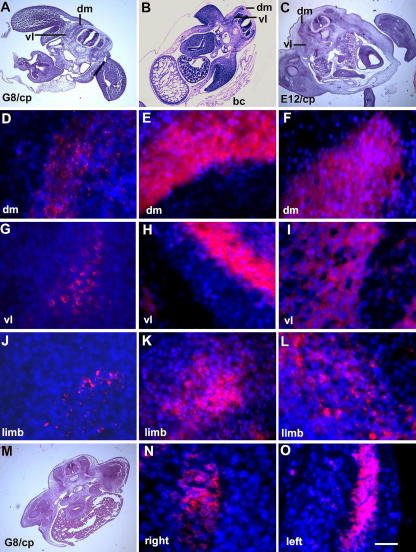
Effect of ablating MyoDpos/G8pos epiblast cells on skeletal muscle differentiation in the somites and limbs. Stage 2 G8pos epiblast cells were lysed with complement (G8/cp). Controls included embryos treated with E12 and complement (E12/cp) or Hanks' buffer only (bc). Embryos were incubated for 4.5–5 d (stages 25–26). Regions on the right side of the embryo in hematoxylin and eosin–stained sections (A–C) are shown at higher magnification in fluorescence photomicrographs of merged images of Hoechst-stained nuclei and rhodamine-labeled antibodies to sarcomeric myosin. Less myosin was detected in the dorsomedial (dm) and ventrolateral (vl) portions of the myotome and limb of G8/complement-treated embryos (D, G, and J) than in those treated with buffer (E, H, and K) or E12 mAb/complement (F, I, and L). (M) A hematoxylin and eosin–stained section shown at higher magnification in N and O. Myosin expression was affected more severely on the right side of the embryo than on the left. Bar: (A–C and M) 135 μm; (N and O) 9 μm in D–L.
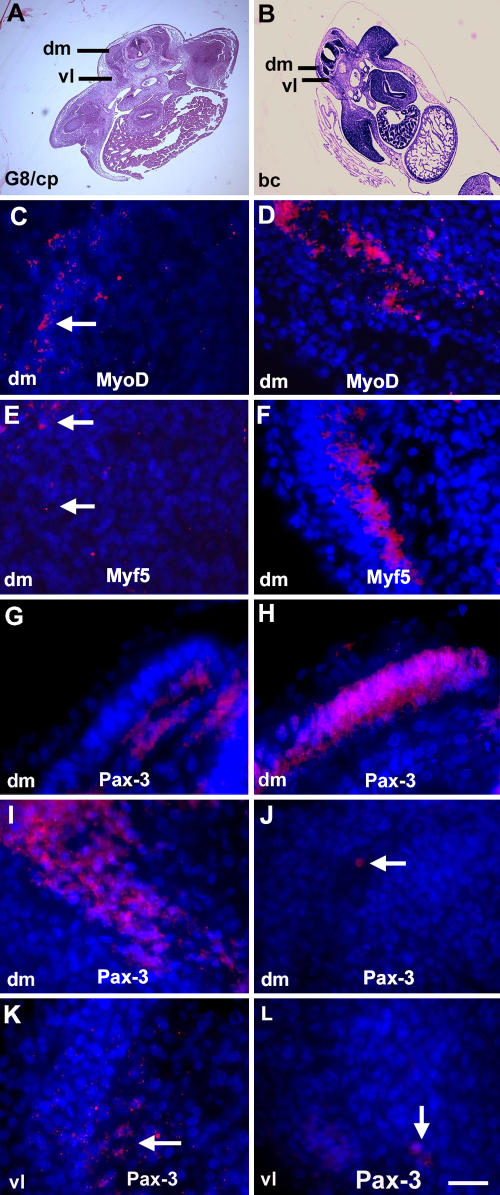
Effect of ablating MyoDpos/G8pos epiblast cells on MyoD, Myf5, and Pax-3 expression. Stage 2 embryos were treated with the G8 mAb and complement (G8/cp) or Hanks' buffer (bc) and grown for 2 d (stage 14; G and H) or 4.5 d (stage 25; C–F and I–L). Areas outlined in hematoxylin and eosin–stained sections (A and B) are shown at higher magnification in fluorescence photomicrographs that are the merged images of Hoechst-stained nuclei and Cy3-labeled dendrimers to MyoD or Myf5 mRNA, or rhodamine-labeled antibody to Pax-3. Less MyoD and Myf5 mRNAs were detected in the somites of G8/complement-treated embryos (C and E) than in embryos treated with buffer only (D and F). More Pax-3–positive cells were present in stage 14 control embryos than treated embryos (G and H), whereas the reverse was found in dorsomedial (dm) and ventrolateral (vl) dermomyotomes/myotomes of stage 25 embryos (I–L). Bar: (A and B) 135 μm; (C–L) 9 μm.
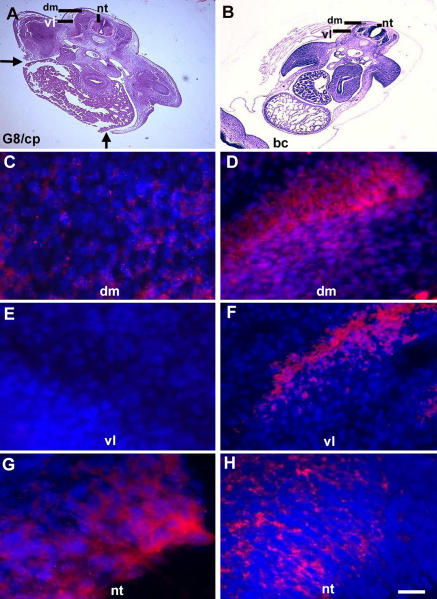
Effect of ablating G8pos/MyoDpos epiblast cells on Noggin expression. Noggin expression was analyzed 4.5 d after ablating stage 2 MyoDpos epiblast cells. Regions indicated in hematoxylin and eosin–stained sections (A and B) are shown in fluorescence photomicrographs of merged images of Hoechst-stained nuclei and Cy3-labeled dendrimers to Noggin mRNA (C–H). Less Noggin mRNA was detected in the dorsomedial (dm) and ventrolateral (vl) dermomyotome and myotome in G8/complement-treated embryos (G8/cp) (C and E) than in buffer-treated embryos (bc; D and F). (G and H) The reverse was found for Noggin expression in the neural tube (nt). Bar: (A and B) 135 μm; (C–H) 9 μm.
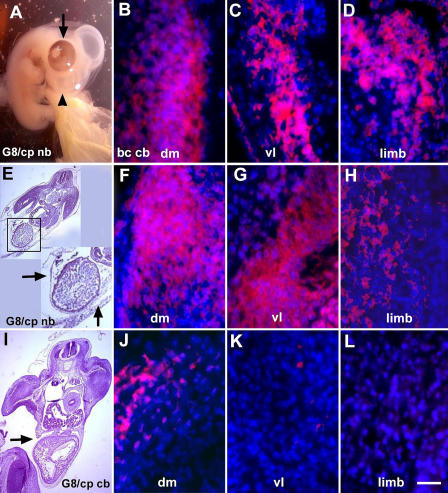
Effect of exogenous Noggin on muscle differentiation in G8/complement-treated embryos. Noggin-soaked beads were implanted lateral to the somites 2 d after ablating MyoDpos/G8pos cells in the stage 2 epiblast. Exogenous Noggin prevented body wall herniations (arrows in inset of E) and facial defects (arrowhead in A), and promoted normal eye pigmentation (arrow in A) and myosin synthesis in the myotome (dm/vl) and limb (F–H). The body wall remained incomplete (arrow in I) and myosin staining was reduced (J–L) in G8/complement-treated embryos implanted with PBS-soaked beads. Staining for myosin in buffer-treated embryos implanted with PBS-soaked beads is shown in B–D. Bar: (A, E, and I) 135 μm; (B–D, F–H, and J–L) 9 μm.
References
-
- Amthor, H., B. Christ, and K. Patel. 1999. A molecular mechanism enabling continuous embryonic muscle growth-a balance between proliferation and differentiation. Development. 126:1041–1053. - PubMed
-
- Baker, C.V.H., M.R. Stark, C. Marcelle, and M. Bronner-Fraser. 1999. Competence, specification and induction of Pax-3 in the trigeminal placode. Development. 126:147–156. - PubMed
-
- Bate, M. 1990. The embryonic development of larval muscles in Drosophila. Development. 110:791–804. - PubMed
-
- Baylies, M.K., and A.M. Michelson. 2001. Invertebrate myogenesis: looking back to the future of muscle development. Curr. Opin. Genet. Dev. 11:431–439. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
