

Crystal structure of a photoactivated deprotonated intermediate of rhodopsin
- PMID: 17060607
- PMCID: PMC1637547
- DOI: 10.1073/pnas.0608022103
Crystal structure of a photoactivated deprotonated intermediate of rhodopsin
Abstract
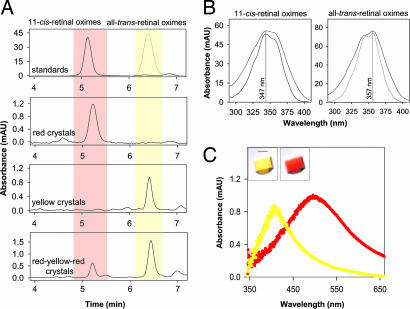
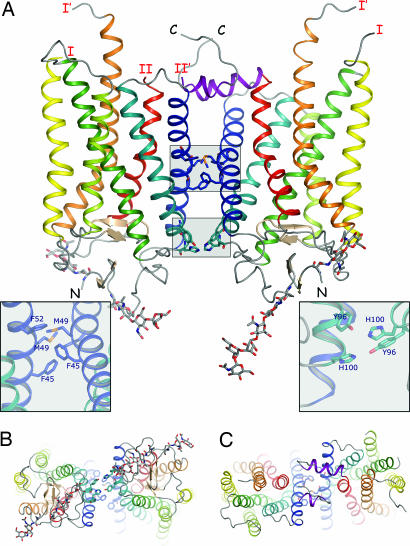
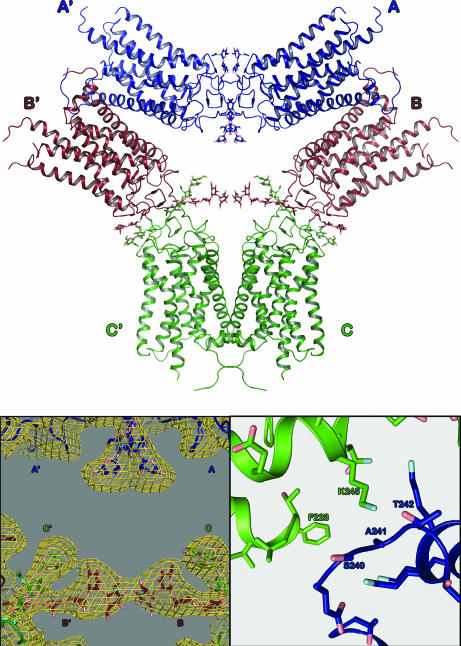
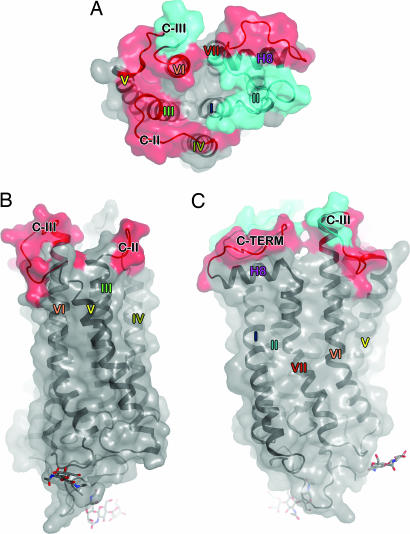
The changes that lead to activation of G protein-coupled receptors have not been elucidated at the structural level. In this work we report the crystal structures of both ground state and a photoactivated deprotonated intermediate of bovine rhodopsin at a resolution of 4.15 A. In the photoactivated state, the Schiff base linking the chromophore and Lys-296 becomes deprotonated, reminiscent of the G protein-activating state, metarhodopsin II. The structures reveal that the changes that accompany photoactivation are smaller than previously predicted for the metarhodopsin II state and include changes on the cytoplasmic surface of rhodopsin that possibly enable the coupling to its cognate G protein, transducin. Furthermore, rhodopsin forms a potentially physiologically relevant dimer interface that involves helices I, II, and 8, and when taken with the prior work that implicates helices IV and V as the physiological dimer interface may account for one of the interfaces of the oligomeric structure of rhodopsin seen in the membrane by atomic force microscopy. The activation and oligomerization models likely extend to the majority of other G protein-coupled receptors.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Foord SM, Bonner TI, Neubig RR, Rosser EM, Pin JP, Davenport AP, Spedding M, Harmar AJ. Pharmacol Rev. 2005;57:279–288. - PubMed
-
- Palczewski K, Kumasaka T, Hori T, Behnke CA, Motoshima H, Fox BA, Le Trong I, Teller DC, Okada T, Stenkamp RE, et al. Science. 2000;289:739–745. - PubMed
-
- Li J, Edwards PC, Burghammer M, Villa C, Schertler GF. J Mol Biol. 2004;343:1409–1438. - PubMed
-
- Okada T, Sugihara M, Bondar AN, Elstner M, Entel P, Buss V. J Mol Biol. 2004;342:571–583. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
