

Murine inner cell mass-derived lineages depend on Sall4 function
- PMID: 17060609
- PMCID: PMC1637580
- DOI: 10.1073/pnas.0607884103
Murine inner cell mass-derived lineages depend on Sall4 function
Abstract
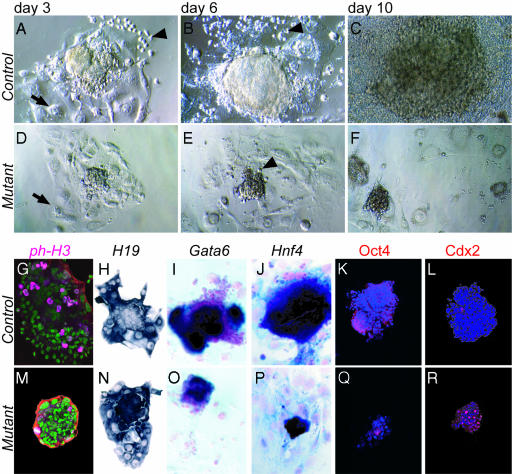
Sall4 is a mammalian Spalt transcription factor expressed by cells of the early embryo and germ cells, an expression pattern similar to that of both Oct4 and Sox2, which play essential roles during early murine development. We show that the activity of Sall4 is cell-autonomously required for the development of the epiblast and primitive endoderm from the inner cell mass. Furthermore, no embryonic or extraembryonic endoderm stem cell lines could be established from Sall4-deficient blastocysts. In contrast, neither the development of the trophoblast lineage nor the ability to generate trophoblast cell lines from murine blastocysts was impaired in the absence of Sall4. These data establish Sall4 as an essential transcription factor required for the early development of inner cell mass-derived cell lineages.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Beddington RS, Robertson EJ. Cell. 1999;96:195–209. - PubMed
-
- Rossant J, Tam PP. Dev Cell. 2004;7:155–164. - PubMed
-
- Johnson MH, McConnell JM. Semin Cell Dev Biol. 2004;15:583–597. - PubMed
-
- Chazaud C, Yamanaka Y, Pawson T, Rossant J. Dev Cell. 2006;10:615–624. - PubMed
-
- Kunath T, Arnaud D, Uy GD, Okamoto I, Chureau C, Yamanaka Y, Heard E, Gardner RL, Avner P, Rossant J. Development (Cambridge, UK) 2005;132:1649–1661. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
