

Function, ontogeny and canalization of shape variance in the primate scapula
- PMID: 17062020
- PMCID: PMC2100339
- DOI: 10.1111/j.1469-7580.2006.00639.x
Function, ontogeny and canalization of shape variance in the primate scapula
Abstract
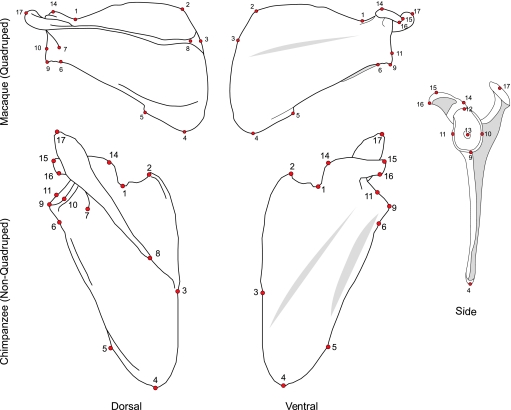
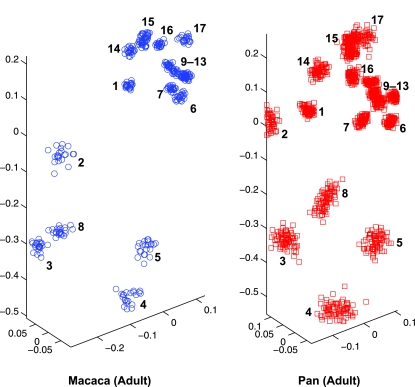
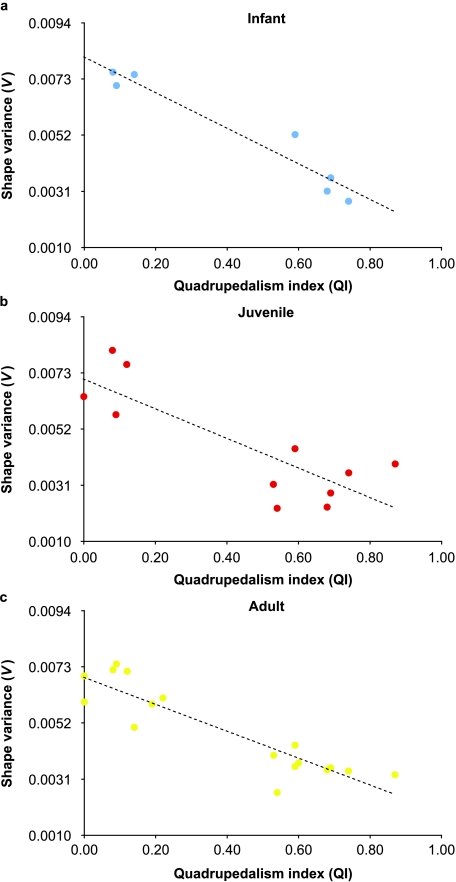
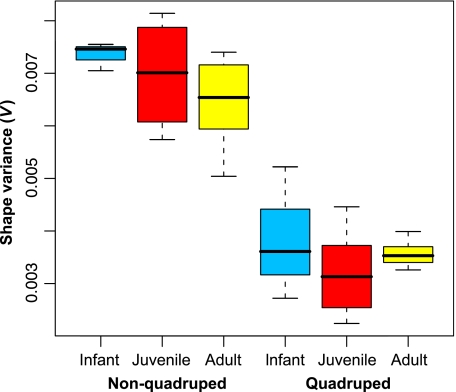
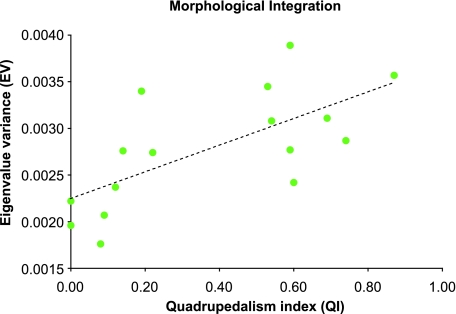
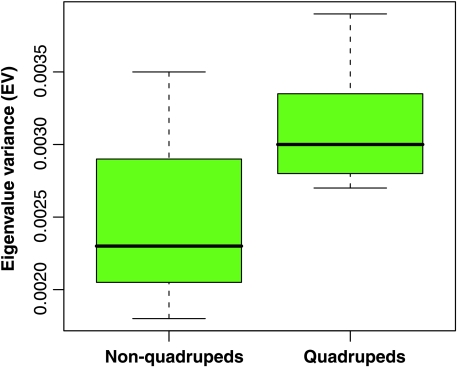
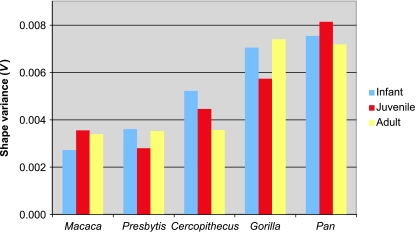
Primates have shoulders adapted to a wide range of locomotor functions from terrestrial pronograde quadrupedalism to highly arboreal suspensory behaviours. The shape of the scapula tightly follows these functional differences. Previous analyses of primate postcrania, including the scapula, indicate that quadrupedal monkeys are less variable than non-quadrupeds. It was previously suggested that this difference was due to a relationship between the strength of stabilizing selection and the functional demands of the upper limb. Here it is shown that intraspecific scapular shape variance is highly correlated with the degree of committed quadrupedalism. Primates that engage in frequent suspensory behaviours (e.g. apes and ateline monkeys) average twice the amount of shape variance as quadrupeds (e.g. Old World monkeys and Saimiri). Because this difference in intraspecific shape variance is apparent in infants and does not increase or decrease appreciably over ontogeny, it is not likely that differences in postnatal growth, neuromuscular control or environmental factors such as habitat structure/composition are the primary contributors to differences in adult shape variance. Instead variance in embryonic factors that affect the shape/size of the scapula or epigenetic factors associated with muscle attachments are more likely candidates. In particular, the heterogeneous functional demands of the non-quadrupedal shoulder probably reduce the stringency of stabilizing selection, resulting in the persistence into adulthood of increased amounts of embryonically generated scapular shape variance.
Figures
Similar articles
-
A comparison of the ontogeny of shape variation in the anthropoid scapula: functional and phylogenetic signal.Am J Phys Anthropol. 2008 Jul;136(3):247-64. doi: 10.1002/ajpa.20799. Am J Phys Anthropol. 2008. PMID: 18324623
-
Scapular position in primates.Folia Primatol (Basel). 2007;78(1):19-35. doi: 10.1159/000095683. Folia Primatol (Basel). 2007. PMID: 17170554
-
Functional morphology of the douc langur (Pygathrix spp.) scapula.Am J Primatol. 2017 Jun;79(6). doi: 10.1002/ajp.22646. Epub 2017 Mar 3. Am J Primatol. 2017. PMID: 28257561
-
Developments in development: What have we learned from primate locomotor ontogeny?Am J Phys Anthropol. 2018 Jan;165 Suppl 65:37-71. doi: 10.1002/ajpa.23388. Am J Phys Anthropol. 2018. PMID: 29380887 Review.
-
Primate quadrupedalism: how and why does it differ from that of typical quadrupeds?Brain Behav Evol. 1989;34(6):357-64. doi: 10.1159/000116522. Brain Behav Evol. 1989. PMID: 2692779 Review.
Cited by
-
Three-dimensional scapular morphology is associated with rotator cuff tears and alters the abduction moment arm of the supraspinatus.Clin Biomech (Bristol). 2020 Aug;78:105091. doi: 10.1016/j.clinbiomech.2020.105091. Epub 2020 Jun 11. Clin Biomech (Bristol). 2020. PMID: 32580097 Free PMC article.
-
Development and the evolvability of human limbs.Proc Natl Acad Sci U S A. 2010 Feb 23;107(8):3400-5. doi: 10.1073/pnas.0911856107. Epub 2010 Feb 1. Proc Natl Acad Sci U S A. 2010. PMID: 20133636 Free PMC article.
-
3D Morphometric and posture study of felid scapulae using statistical shape modelling.PLoS One. 2012;7(4):e34619. doi: 10.1371/journal.pone.0034619. Epub 2012 Apr 11. PLoS One. 2012. PMID: 22509335 Free PMC article.
-
The modular organization of roe deer (Capreolus capreolus) body during ontogeny: the effects of sex and habitat.Front Zool. 2018 Sep 27;15:37. doi: 10.1186/s12983-018-0283-8. eCollection 2018. Front Zool. 2018. PMID: 30275869 Free PMC article.
-
A comparative probabilistic analysis of human and chimpanzee rotator cuff functional capacity.J Anat. 2023 Sep;243(3):431-447. doi: 10.1111/joa.13882. Epub 2023 Apr 25. J Anat. 2023. PMID: 37186281 Free PMC article.
References
-
- Ashton EH, Oxnard CE. The musculature of the primate shoulder. Trans Zool Soc Lond. 1963;29:554–650.
-
- Ashton EH, Flinn RM, Oxnard CE, Spence TF. The adaptive and classificatory significance of certain quantitative features of the forelimb in primates. J Zool (Lond) 1976;179:515–556.
-
- Atchley WR, Hall BK. A model for development and evolution of complex morphological structures. Biol Rev. 1991;66:101–157. - PubMed
-
- Boinski S. The positional behavior and substrate use of squirrel monkeys: ecological implications. J Hum Evol. 1989;18:659–677.
-
- Cant JGH. Positional behavior of long-tailed macaques (Macaca fasicularis) in Northern Sumatra. Am J Phys Anthropol. 1988;76:29–37. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
