

The Slc35d3 gene, encoding an orphan nucleotide sugar transporter, regulates platelet-dense granules
- PMID: 17062724
- PMCID: PMC1794067
- DOI: 10.1182/blood-2006-08-040196
The Slc35d3 gene, encoding an orphan nucleotide sugar transporter, regulates platelet-dense granules
Abstract
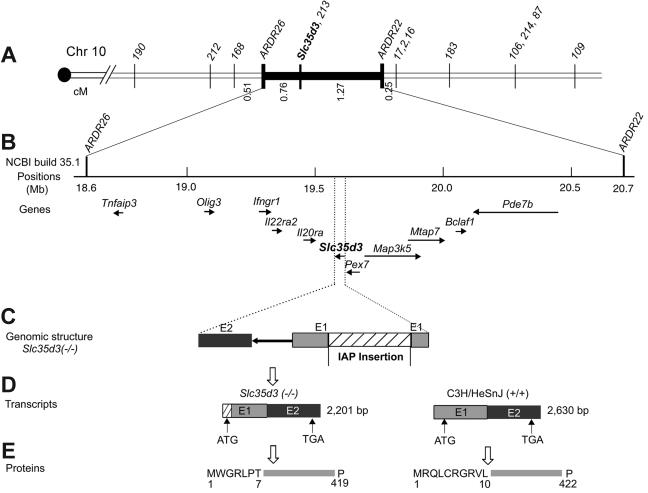
Platelet dense granules are lysosome-related organelles which contain high concentrations of several biologically important low-molecular-weight molecules. These include calcium, serotonin, adenine nucleotides, pyrophosphate, and polyphosphate, which are necessary for normal blood hemostasis. The synthesis of dense granules and other lysosome-related organelles is defective in inherited diseases such as Hermansky-Pudlak syndrome (HPS) and Chediak-Higashi syndrome (CHS). HPS and CHS mutations in 8 human and at least 16 murine genes have been identified. Previous studies produced contradictory findings for the function of the murine ashen (Rab27a) gene in platelet-dense granules. We have used a positional cloning approach with one line of ashen mutants to establish that a new mutation in a second gene, Slc35d3, on mouse chromosome 10 is the basis of this discrepancy. The platelet-dense granule defect is rescued in BAC transgenic mice containing the normal Slc35d3 gene. Thus, Slc35d3, an orphan member of a nucleotide sugar transporter family, specifically regulates the contents of platelet-dense granules. Unlike HPS or CHS genes, it has no apparent effect on other lysosome-related organelles such as melanosomes or lysosomes. The ash-Roswell mouse mutant is an appropriate model for human congenital-isolated delta-storage pool deficiency.
Figures





References
-
- Rendu F, Brohard-Bohn B. The platelet release reaction: granules' constituents, secretion and functions. Platelets. 2001;12:261–273. - PubMed
-
- McNicol A, Israels SJ. Platelet dense granules: structure, function and implications for haemostasis. Thromb Res. 1999;95:1–18. - PubMed
-
- Ruiz FA, Lea CR, Oldfield E, Docampo R. Human platelet-dense granules contain polyphosphate and are similar to acidocalcisomes of bacteria and unicellular eukaryotes. J Biol Chem. 2004;279:44250–44257. - PubMed
-
- Youssefian T, Cramer EM. Megakaryocyte dense granule components are sorted in multivesicular bodies. Blood. 2000;95:4004–4007. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
