

A cryptochrome/photolyase class of enzymes with single-stranded DNA-specific photolyase activity
- PMID: 17062752
- PMCID: PMC1621107
- DOI: 10.1073/pnas.0607993103
A cryptochrome/photolyase class of enzymes with single-stranded DNA-specific photolyase activity
Abstract
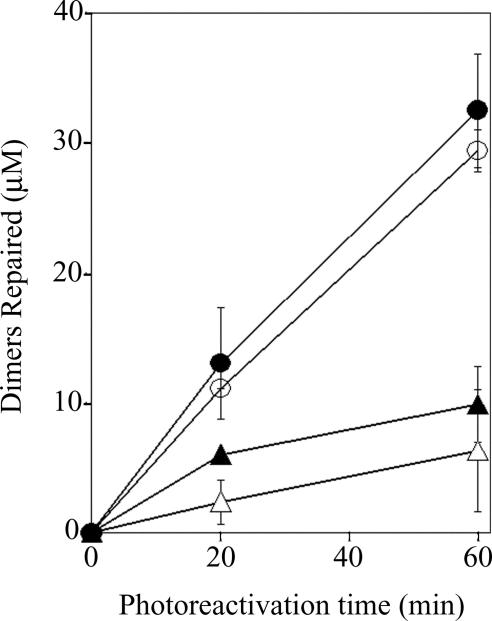
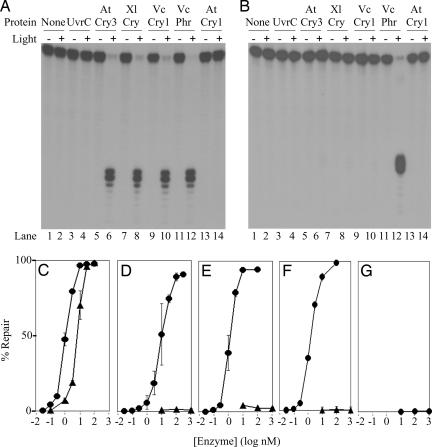
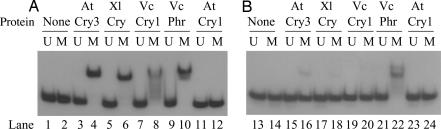
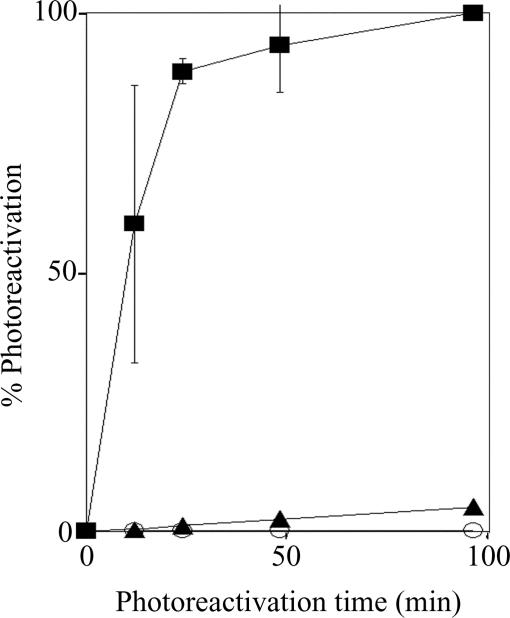
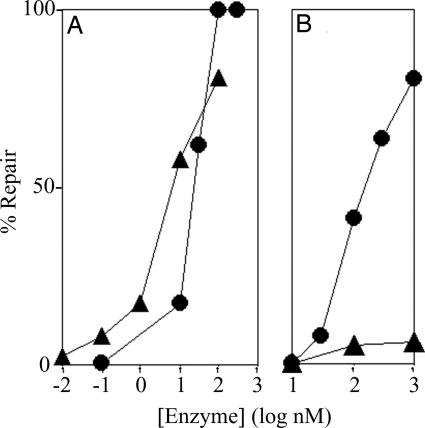
Photolyases and cryptochrome blue-light photoreceptors are evolutionarily related flavoproteins that perform distinct functions. Photolyases repair UV-damaged DNA in many species from bacteria to plants and animals. Cryptochromes regulate growth and development in plants and the circadian clock in animals. Recently, a new branch of the photolyase/cryptochrome family was identified. Members of this branch exhibited no or trace levels of DNA repair activity in vivo and in vitro and, therefore, were considered to be cryptochromes, and they were named cryptochrome-DASH. Here, we show that Cry-DASH proteins from bacterial, plant, and animal sources actually are photolyases with high degree of specificity for cyclobutane pyrimidine dimers in ssDNA.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
