

Structural and mutagenic analysis of foot-and-mouth disease virus 3C protease reveals the role of the beta-ribbon in proteolysis
- PMID: 17065215
- PMCID: PMC1797255
- DOI: 10.1128/JVI.01587-06
Structural and mutagenic analysis of foot-and-mouth disease virus 3C protease reveals the role of the beta-ribbon in proteolysis
Abstract
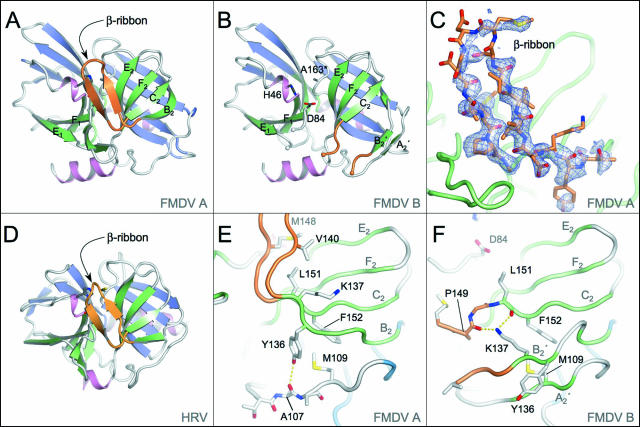
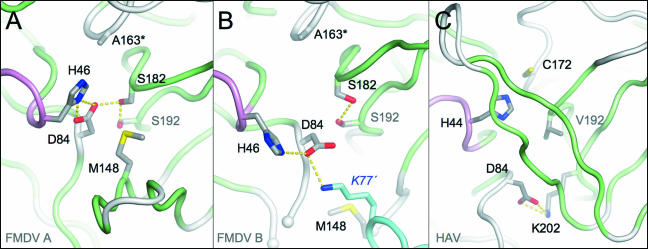
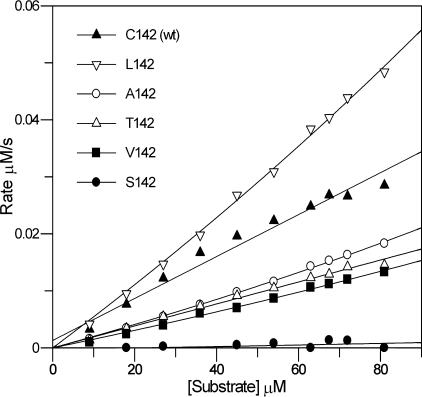
The 3C protease (3C(pro)) from foot-and-mouth disease virus (FMDV), the causative agent of a widespread and economically devastating disease of domestic livestock, is a potential target for antiviral drug design. We have determined the structure of a new crystal form of FMDV 3C(pro), a chymotrypsin-like cysteine protease, which reveals features that are important for catalytic activity. In particular, we show that a surface loop which was disordered in previous structures adopts a beta-ribbon structure that is conformationally similar to equivalent regions on other picornaviral 3C proteases and some serine proteases. This beta-ribbon folds over the peptide binding cleft and clearly contributes to substrate recognition. Replacement of Cys142 at the tip of the beta-ribbon with different amino acids has a significant impact on enzyme activity and shows that higher activity is obtained with more hydrophobic side chains. Comparison of the structure of FMDV 3C(pro) with homologous enzyme-peptide complexes suggests that this correlation arises because the side chain of Cys142 contacts the hydrophobic portions of the P2 and P4 residues in the peptide substrate. Collectively, these findings provide compelling evidence for the role of the beta-ribbon in catalytic activity and provide valuable insights for the design of FMDV 3C(pro) inhibitors.
Figures
Similar articles
-
Insights into cleavage specificity from the crystal structure of foot-and-mouth disease virus 3C protease complexed with a peptide substrate.J Mol Biol. 2010 Jan 15;395(2):375-89. doi: 10.1016/j.jmb.2009.10.048. Epub 2009 Oct 31. J Mol Biol. 2010. PMID: 19883658 Free PMC article.
-
Crystal structure of foot-and-mouth disease virus 3C protease. New insights into catalytic mechanism and cleavage specificity.J Biol Chem. 2005 Mar 25;280(12):11520-7. doi: 10.1074/jbc.M413254200. Epub 2005 Jan 14. J Biol Chem. 2005. PMID: 15654079
-
Structural analysis of foot-and-mouth disease virus 3C protease: a viable target for antiviral drugs?Biochem Soc Trans. 2007 Jun;35(Pt 3):594-8. doi: 10.1042/BST0350594. Biochem Soc Trans. 2007. PMID: 17511659 Review.
-
Crystal structure of human enterovirus 71 3C protease.J Mol Biol. 2011 May 6;408(3):449-61. doi: 10.1016/j.jmb.2011.03.007. Epub 2011 Mar 17. J Mol Biol. 2011. PMID: 21396941 Free PMC article.
-
Foot-and-mouth disease virus 3C protease: recent structural and functional insights into an antiviral target.Int J Biochem Cell Biol. 2007;39(1):1-6. doi: 10.1016/j.biocel.2006.07.006. Epub 2006 Aug 14. Int J Biochem Cell Biol. 2007. PMID: 16979372 Free PMC article. Review.
Cited by
-
Cap Is the Protease of the Porcine Circovirus 2.Viruses. 2022 Jul 15;14(7):1550. doi: 10.3390/v14071550. Viruses. 2022. PMID: 35891530 Free PMC article.
-
Genetic heterogeneity in the leader and P1-coding regions of foot-and-mouth disease virus serotypes A and O in Africa.Arch Virol. 2014 May;159(5):947-61. doi: 10.1007/s00705-013-1838-9. Epub 2013 Nov 13. Arch Virol. 2014. PMID: 24221247 Free PMC article.
-
Shifts in the selection-drift balance drive the evolution and epidemiology of foot-and-mouth disease virus.J Virol. 2009 Jan;83(2):781-90. doi: 10.1128/JVI.01500-08. Epub 2008 Nov 12. J Virol. 2009. PMID: 19004952 Free PMC article.
-
Identification and characterization of Iflavirus 3C-like protease processing activities.Virology. 2012 Jul 5;428(2):136-45. doi: 10.1016/j.virol.2012.04.002. Epub 2012 Apr 24. Virology. 2012. PMID: 22534091 Free PMC article.
-
Foot-and-mouth disease virus 3C protease induces fragmentation of the Golgi compartment and blocks intra-Golgi transport.J Virol. 2013 Nov;87(21):11721-9. doi: 10.1128/JVI.01355-13. Epub 2013 Aug 28. J Virol. 2013. PMID: 23986596 Free PMC article.
References
-
- Allaire, M., M. M. Chernaia, B. A. Malcolm, and M. N. James. 1994. Picornaviral 3C cysteine proteinases have a fold similar to chymotrypsin-like serine proteinases. Nature 369:72-76. - PubMed
-
- Atherton, E., and R. C. Sheppard (ed.). 1989. Solid phase peptide synthesis. Oxford University Press, Oxford, United Kingdom.
-
- Bergmann, E. M., M. M. Cherney, J. McKendrick, S. Frormann, C. Luo, B. A. Malcolm, J. C. Vederas, and M. N. James. 1999. Crystal structure of an inhibitor complex of the 3C proteinase from hepatitis A virus (HAV) and implications for the polyprotein processing in HAV. Virology 265:153-163. - PubMed
-
- Birtley, J. R., and S. Curry. 2005. Crystallization of foot-and-mouth disease virus 3C protease: surface mutagenesis and a novel crystal-optimization strategy. Acta Crystallogr. D. 61:646-650. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
