

RanBP2 modulates Cox11 and hexokinase I activities and haploinsufficiency of RanBP2 causes deficits in glucose metabolism
- PMID: 17069463
- PMCID: PMC1626108
- DOI: 10.1371/journal.pgen.0020177
RanBP2 modulates Cox11 and hexokinase I activities and haploinsufficiency of RanBP2 causes deficits in glucose metabolism
Abstract
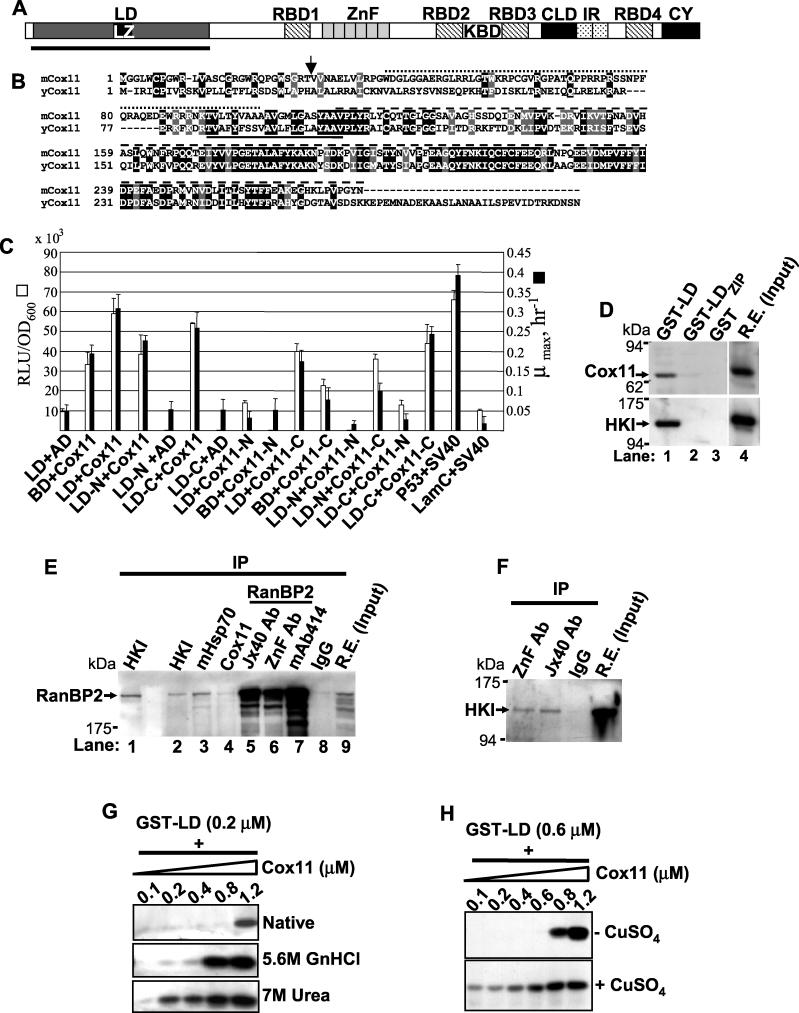
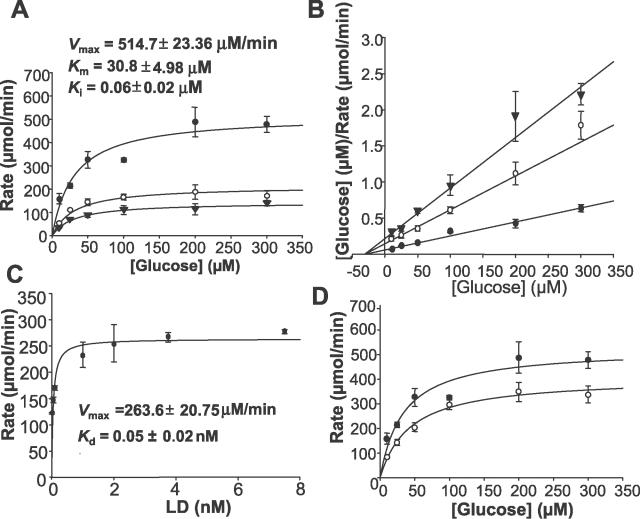
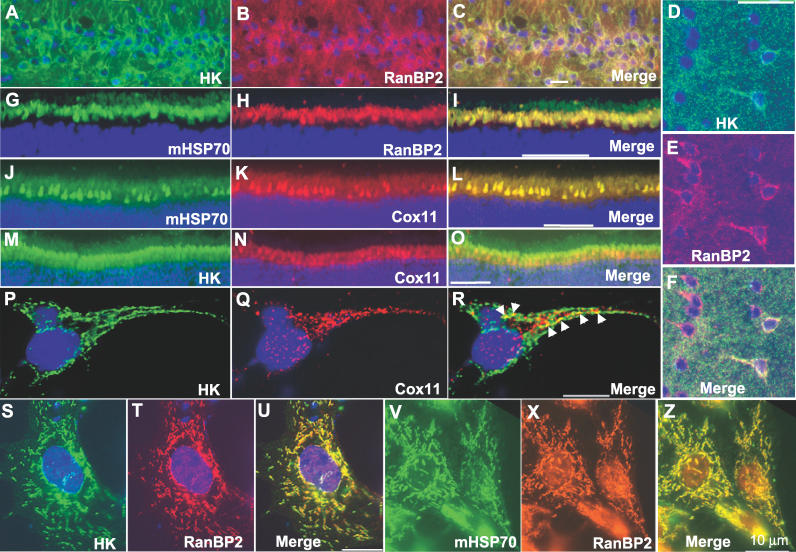
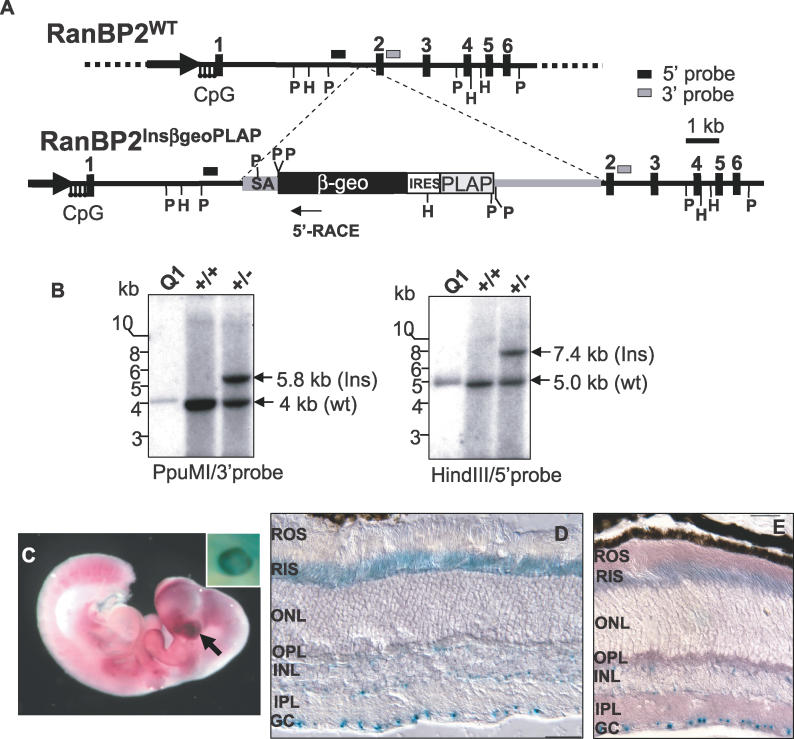
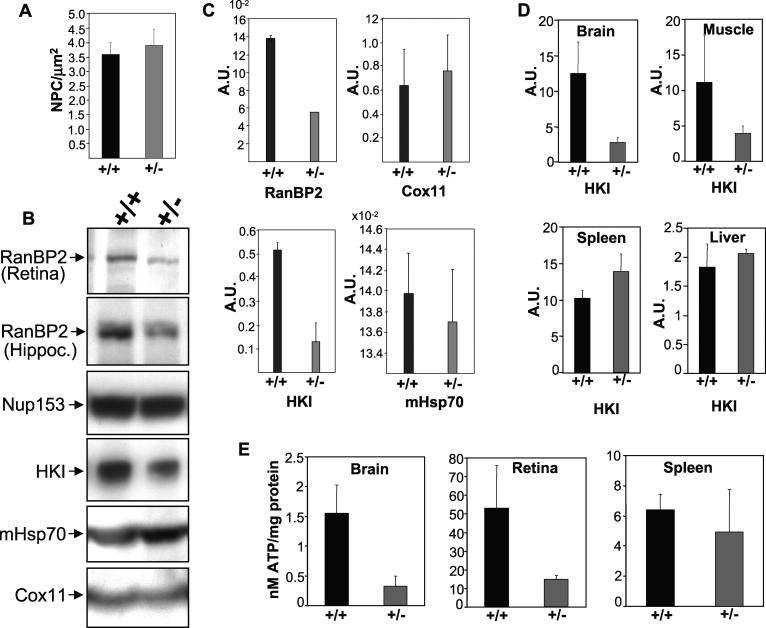
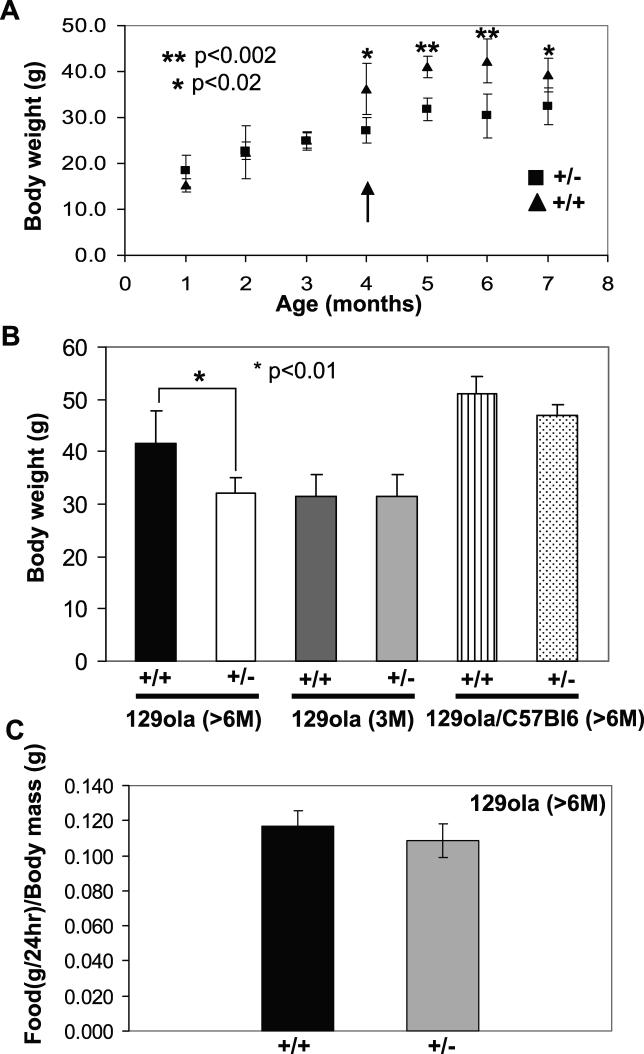
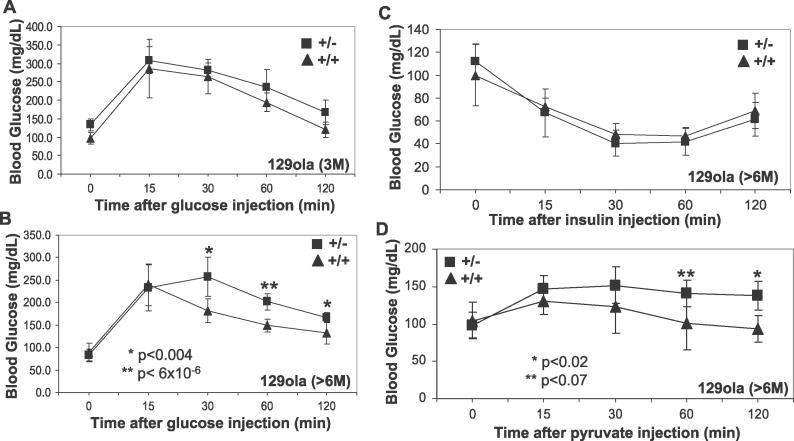
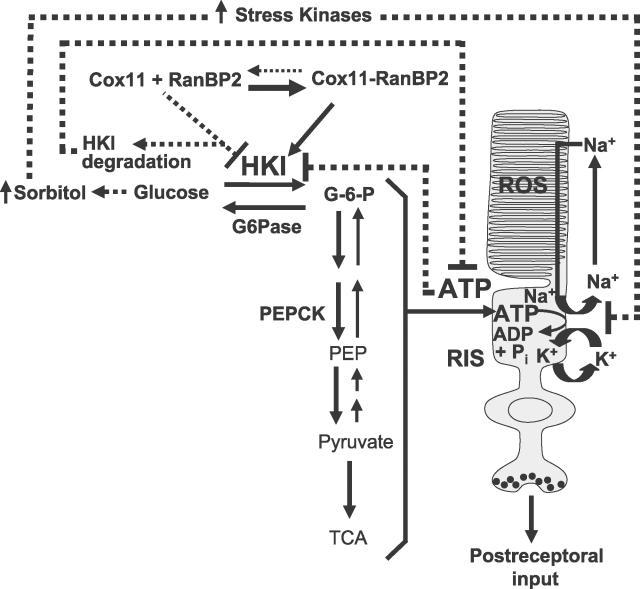
The Ran-binding protein 2 (RanBP2) is a large multimodular and pleiotropic protein. Several molecular partners with distinct functions interacting specifically with selective modules of RanBP2 have been identified. Yet, the significance of these interactions with RanBP2 and the genetic and physiological role(s) of RanBP2 in a whole-animal model remain elusive. Here, we report the identification of two novel partners of RanBP2 and a novel physiological role of RanBP2 in a mouse model. RanBP2 associates in vitro and in vivo and colocalizes with the mitochondrial metallochaperone, Cox11, and the pacemaker of glycolysis, hexokinase type I (HKI) via its leucine-rich domain. The leucine-rich domain of RanBP2 also exhibits strong chaperone activity toward intermediate and mature folding species of Cox11 supporting a chaperone role of RanBP2 in the cytosol during Cox11 biogenesis. Cox11 partially colocalizes with HKI, thus supporting additional and distinct roles in cell function. Cox11 is a strong inhibitor of HKI, and RanBP2 suppresses the inhibitory activity of Cox11 over HKI. To probe the physiological role of RanBP2 and its role in HKI function, a mouse model harboring a genetically disrupted RanBP2 locus was generated. RanBP2(-/-) are embryonically lethal, and haploinsufficiency of RanBP2 in an inbred strain causes a pronounced decrease of HKI and ATP levels selectively in the central nervous system. Inbred RanBP2(+/-) mice also exhibit deficits in growth rates and glucose catabolism without impairment of glucose uptake and gluconeogenesis. These phenotypes are accompanied by a decrease in the electrophysiological responses of photosensory and postreceptoral neurons. Hence, RanBP2 and its partners emerge as critical modulators of neuronal HKI, glucose catabolism, energy homeostasis, and targets for metabolic, aging disorders and allied neuropathies.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures

References
-
- Ferreira PA, Hom JT, Pak WL. Retina-specifically expressed novel subtypes of bovine cyclophilin. J Biol Chem. 1995;270:23179–23188. - PubMed
-
- Wu J, Matunis MJ, Kraemer D, Blobel G, Coutavas E. Nup358, a cytoplasmically exposed nucleoporin with peptide repeats, Ran-GTP binding sites, zinc fingers, a cyclophilin A homologous domain, and a leucine-rich region. J Biol Chem. 1995;270:14209–14213. - PubMed
-
- Yokoyama N, Hayashi N, Seki T, Pante N, Ohba T, et al. A giant nucleopore protein that binds Ran/TC4. Nature. 1995;376:184–188. - PubMed
-
- Fauser S, Aslanukov A, Roepman R, Ferreira PA. Genomic organization, expression, and localization of murine Ran-binding protein 2 (RanBP2) gene. Mamm Genome. 2001;12:406–415. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
