

Characteristics of the nuclear (18S, 5.8S, 28S and 5S) and mitochondrial (12S and 16S) rRNA genes of Apis mellifera (Insecta: Hymenoptera): structure, organization, and retrotransposable elements
- PMID: 17069639
- PMCID: PMC2048585
- DOI: 10.1111/j.1365-2583.2006.00689.x
Characteristics of the nuclear (18S, 5.8S, 28S and 5S) and mitochondrial (12S and 16S) rRNA genes of Apis mellifera (Insecta: Hymenoptera): structure, organization, and retrotransposable elements
Abstract
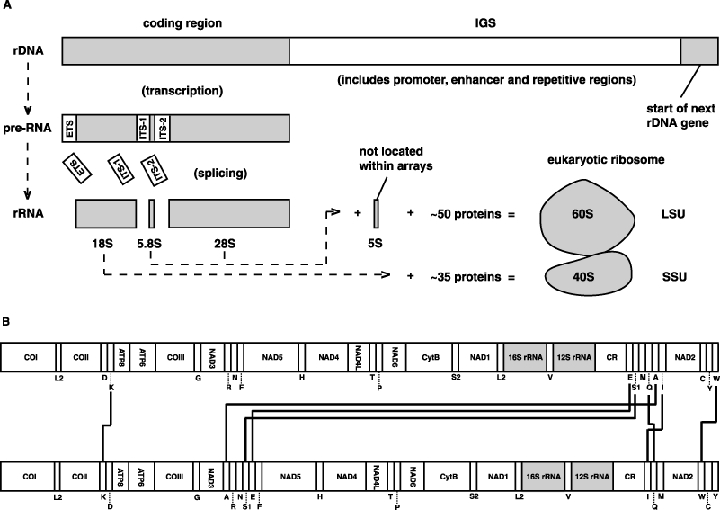
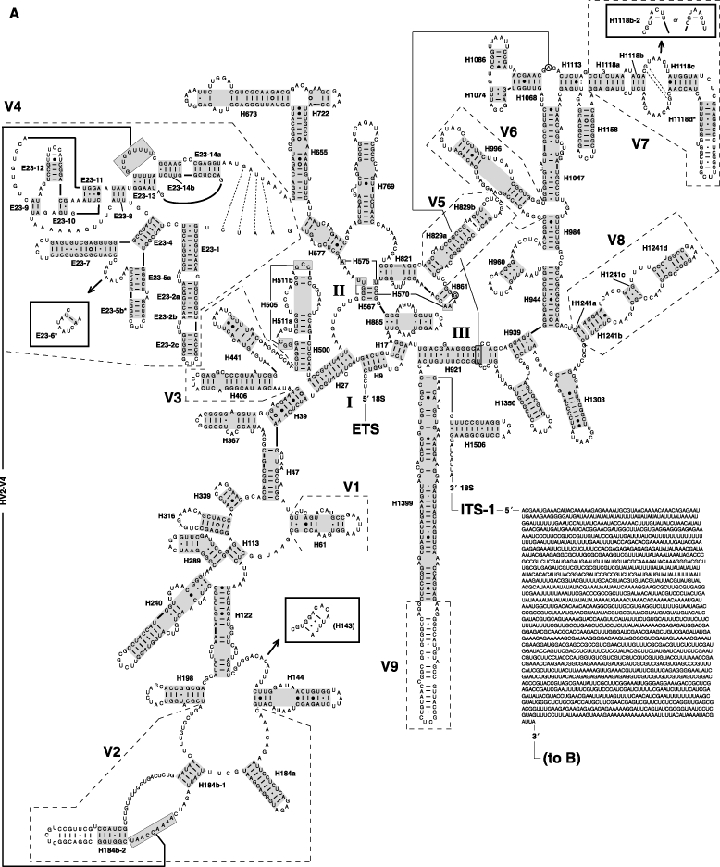
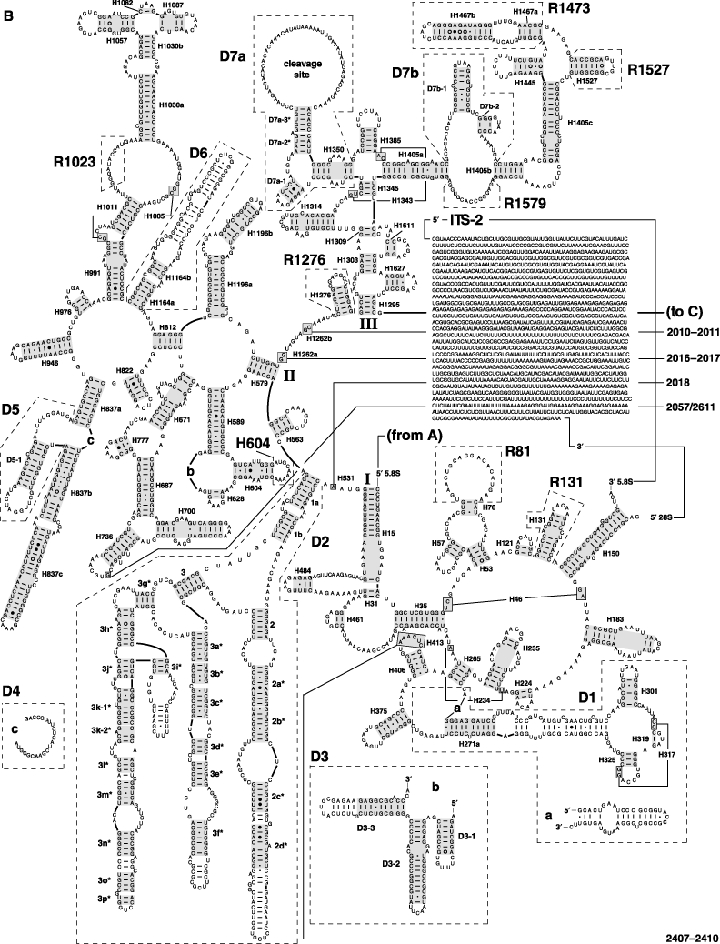
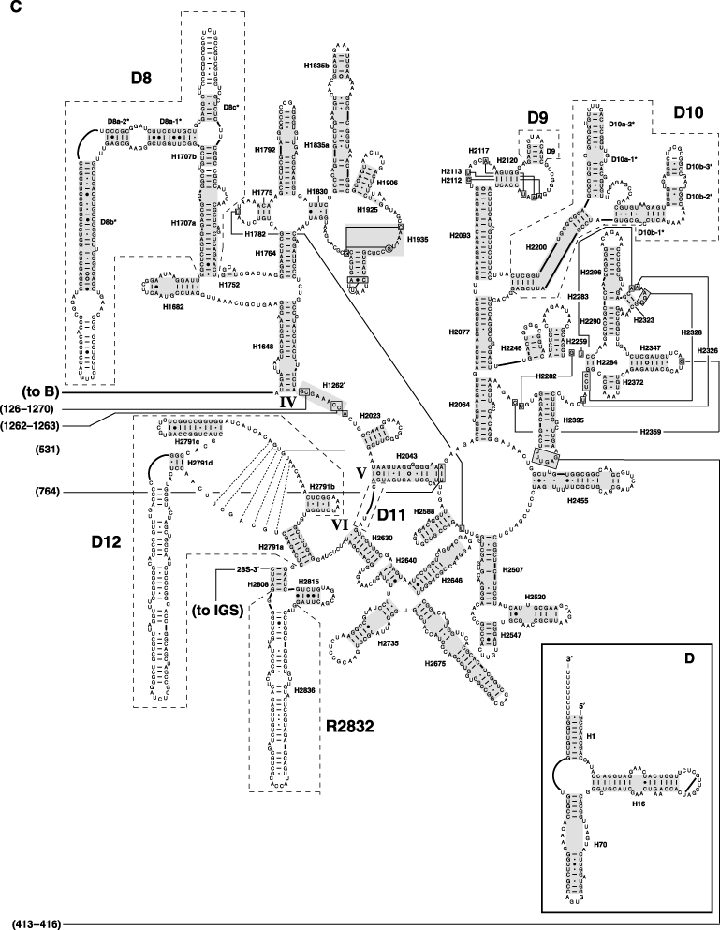
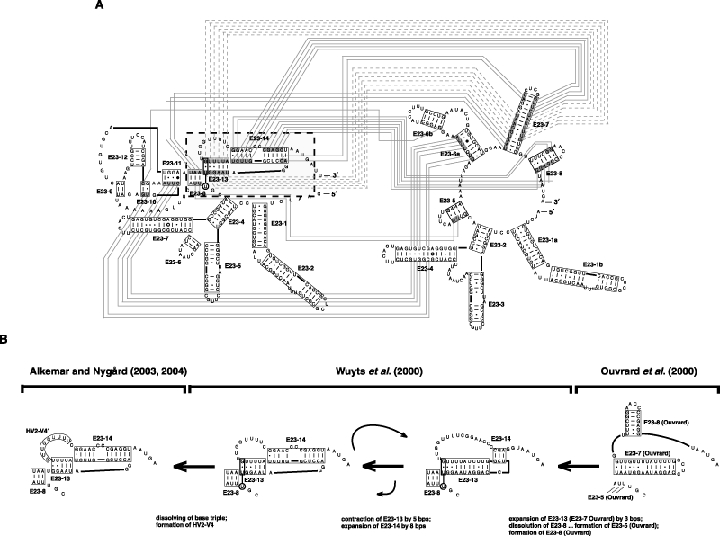
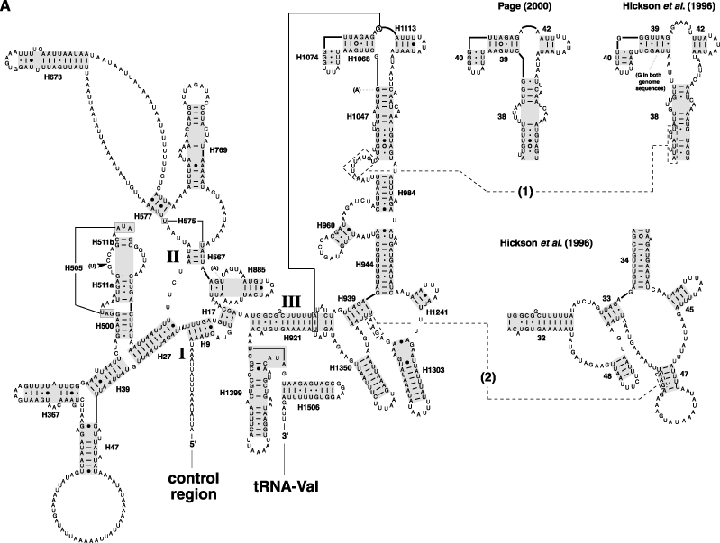
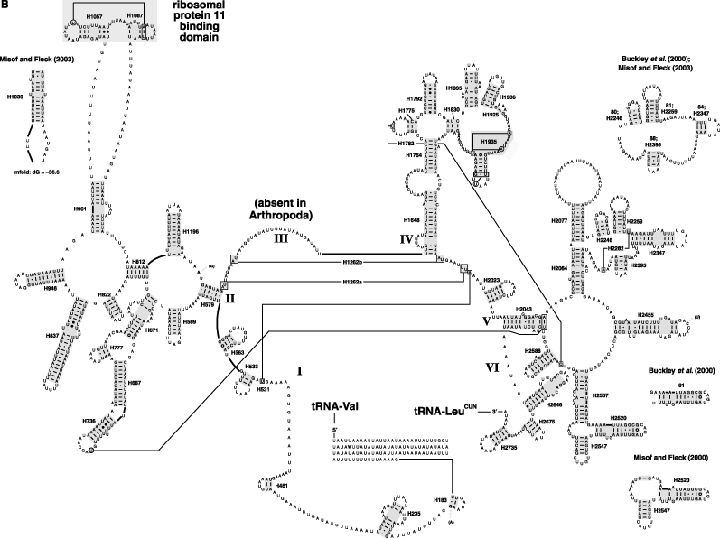
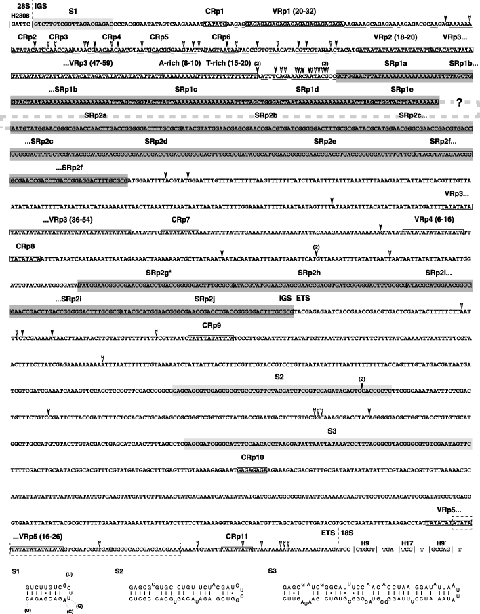
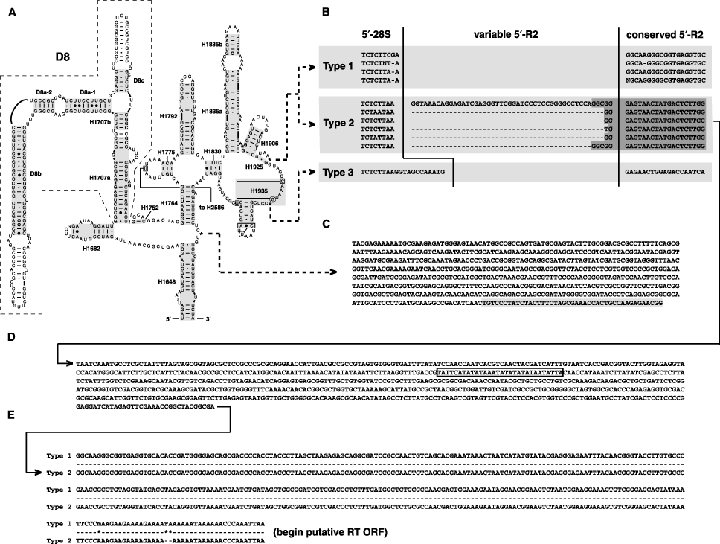
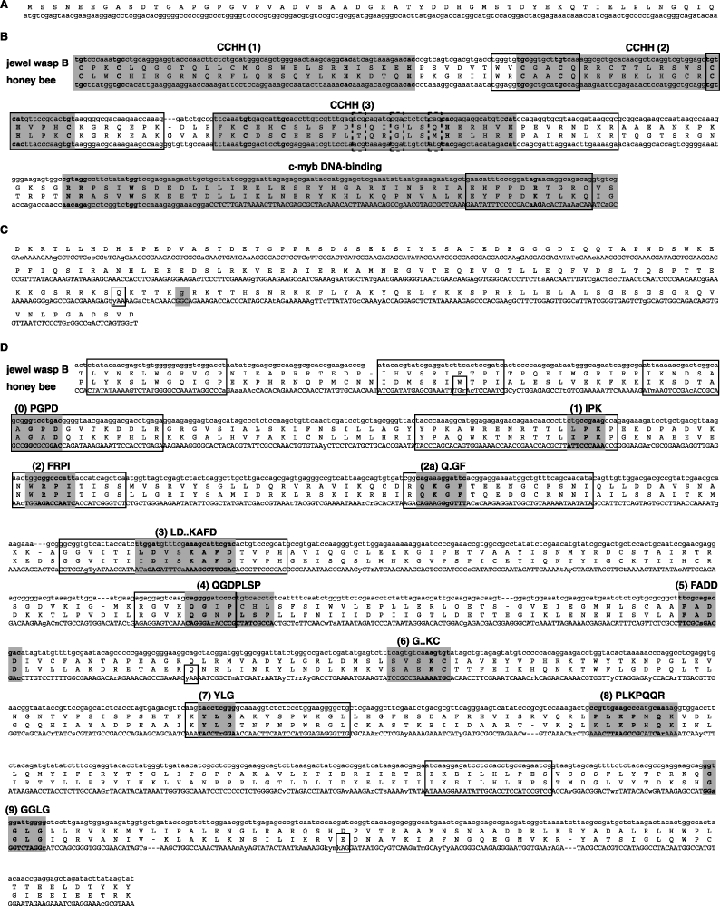
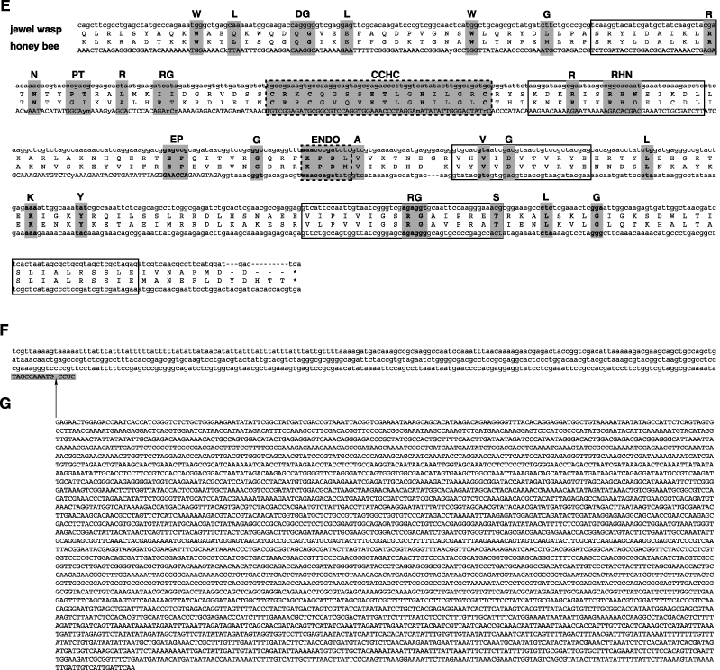
As an accompanying manuscript to the release of the honey bee genome, we report the entire sequence of the nuclear (18S, 5.8S, 28S and 5S) and mitochondrial (12S and 16S) ribosomal RNA (rRNA)-encoding gene sequences (rDNA) and related internally and externally transcribed spacer regions of Apis mellifera (Insecta: Hymenoptera: Apocrita). Additionally, we predict secondary structures for the mature rRNA molecules based on comparative sequence analyses with other arthropod taxa and reference to recently published crystal structures of the ribosome. In general, the structures of honey bee rRNAs are in agreement with previously predicted rRNA models from other arthropods in core regions of the rRNA, with little additional expansion in non-conserved regions. Our multiple sequence alignments are made available on several public databases and provide a preliminary establishment of a global structural model of all rRNAs from the insects. Additionally, we provide conserved stretches of sequences flanking the rDNA cistrons that comprise the externally transcribed spacer regions (ETS) and part of the intergenic spacer region (IGS), including several repetitive motifs. Finally, we report the occurrence of retrotransposition in the nuclear large subunit rDNA, as R2 elements are present in the usual insertion points found in other arthropods. Interestingly, functional R1 elements usually present in the genomes of insects were not detected in the honey bee rRNA genes. The reverse transcriptase products of the R2 elements are deduced from their putative open reading frames and structurally aligned with those from another hymenopteran insect, the jewel wasp Nasonia (Pteromalidae). Stretches of conserved amino acids shared between Apis and Nasonia are illustrated and serve as potential sites for primer design, as target amplicons within these R2 elements may serve as novel phylogenetic markers for Hymenoptera. Given the impending completion of the sequencing of the Nasonia genome, we expect our report eventually to shed light on the evolution of the hymenopteran genome within higher insects, particularly regarding the relative maintenance of conserved rDNA genes, related variable spacer regions and retrotransposable elements.
Figures
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Amako D, Kwon OY, Ishikawa H. Nucleotide sequence and presumed secondary structure of the 28S rRNA of pea aphid: implication for diversification of insect rRNA. J Mol Evol. 1996;206:469–475. - PubMed
-
- Applebaum SW, Ebstein RP, Wyatt GR. Dissociation of ribosomal ribonucleic acid from silkmoth pupae by heat and dimethylsulfoxide: evidence for specific cleavage points. J Mol Biol. 1966;21:29–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
