

Determining the critical nucleus and mechanism of fibril elongation of the Alzheimer's Abeta(1-40) peptide
- PMID: 17070840
- PMCID: PMC2674024
- DOI: 10.1016/j.jmb.2006.10.011
Determining the critical nucleus and mechanism of fibril elongation of the Alzheimer's Abeta(1-40) peptide
Abstract
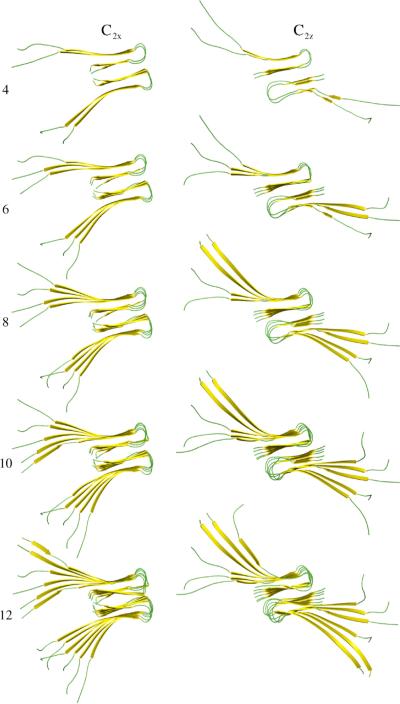
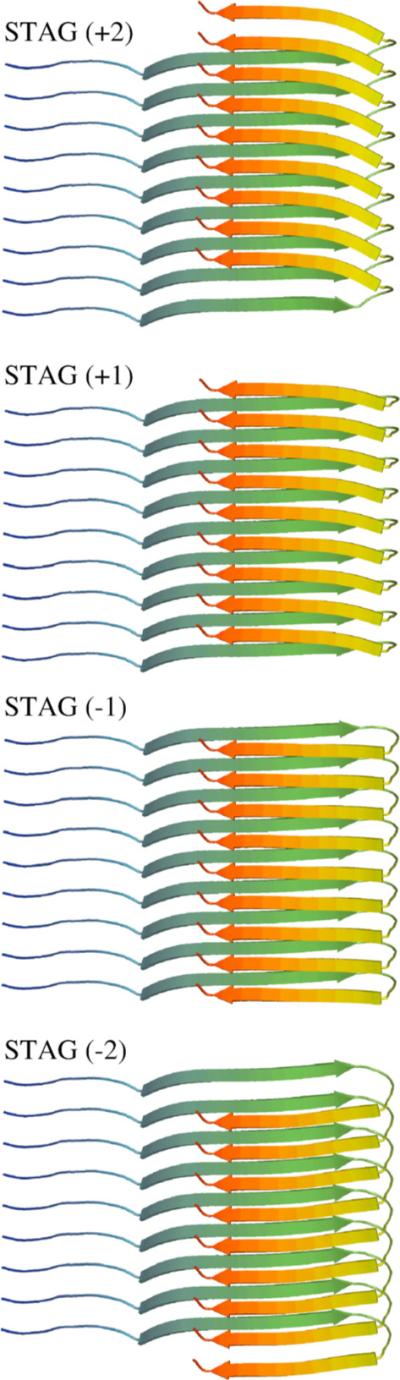
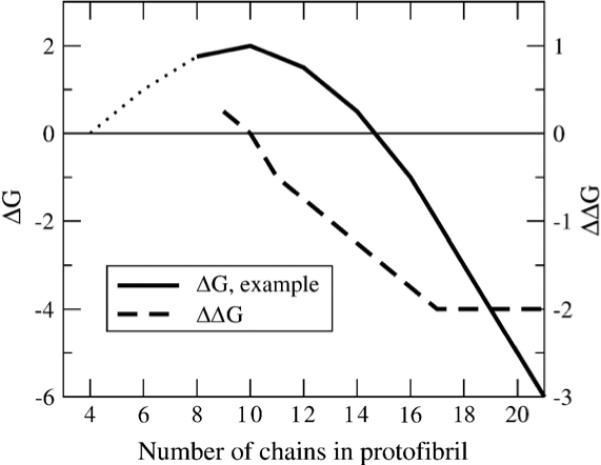
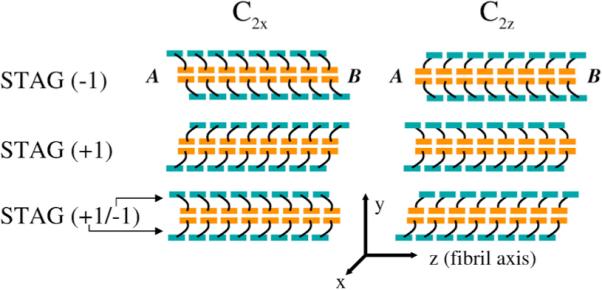
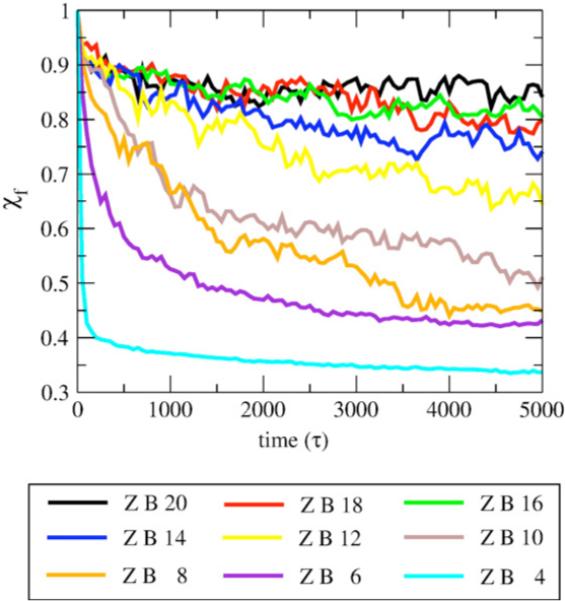
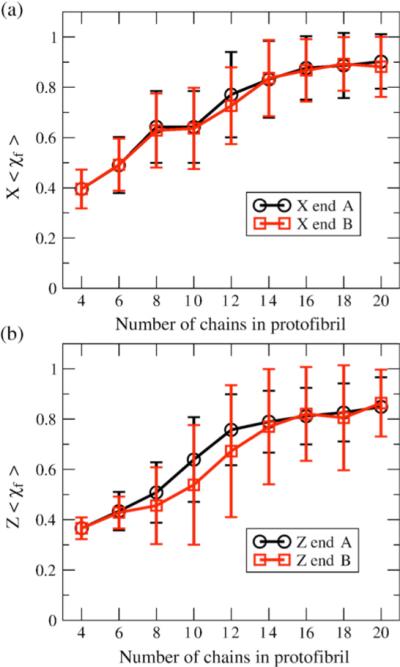
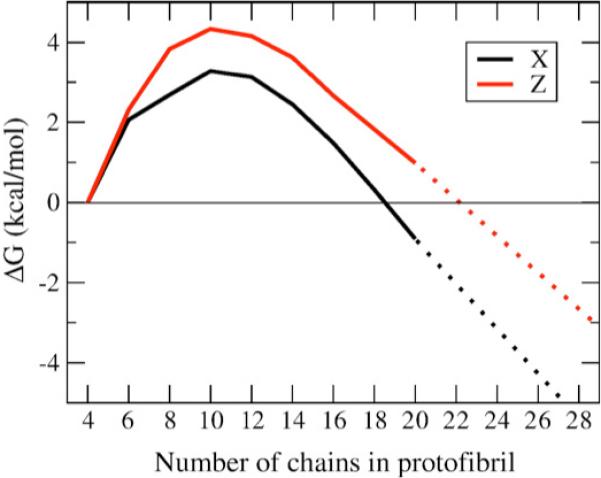
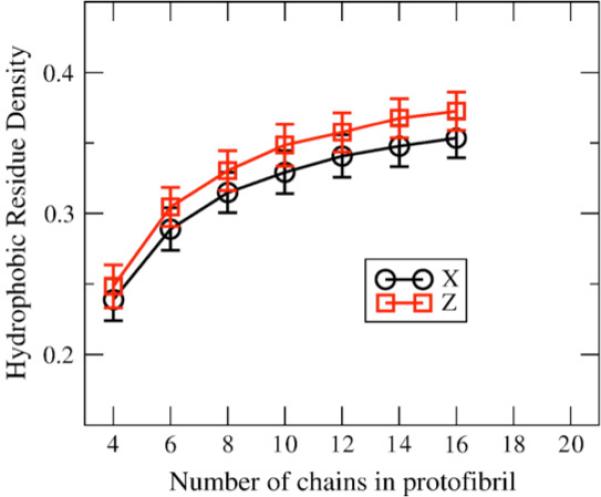
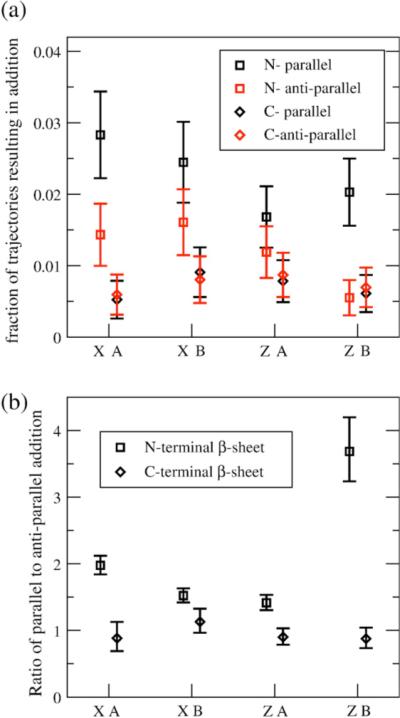
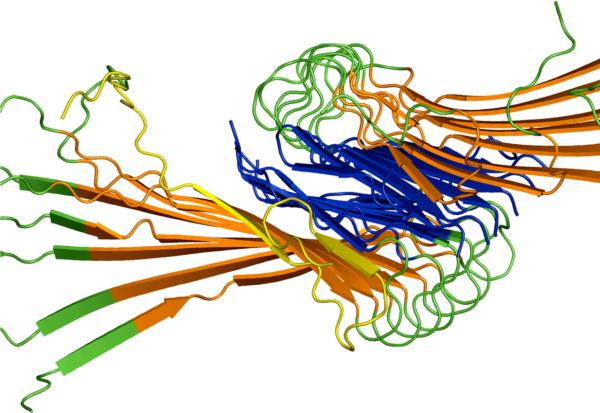
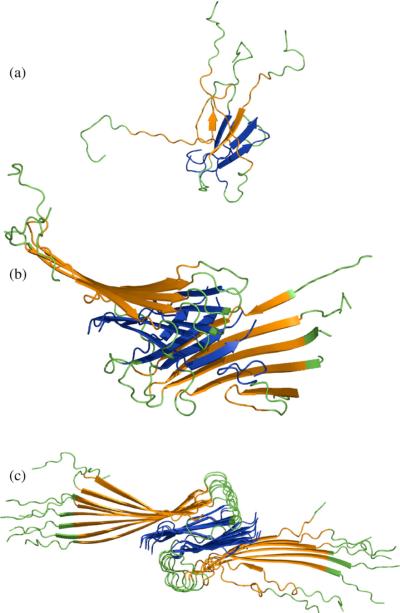
We use a coarse-grained protein model to characterize the critical nucleus, structural stability, and fibril elongation propensity of Abeta(1-40) oligomers for the C(2x) and C(2z) quaternary forms proposed by solid-state NMR. By estimating equilibrium populations of structurally stable and unstable protofibrils, we determine the shift in the dominant population from free monomer to ordered fibril at a critical nucleus of ten chains for the C(2x) and C(2z) forms. We find that a minimum assembly of 16 monomer chains is necessary to mimic a mature fibril, and show that its structural stability correlates with a plateau in the hydrophobic residue density and a decrease in the likelihood of losing hydrophobic interactions by rotating the fibril subunits. While Abeta(1-40) protofibrils show similar structural stability for both C(2x) and C(2z) quaternary structures, we find that the fibril elongation propensity is greater for the C(2z) form relative to the C(2x) form. We attribute the increased propensity for elongation of the C(2z) form as being due to a stagger in the interdigitation of the N-terminal and C-terminal beta-strands, resulting in structural asymmetry in the presented fibril ends that decreases the amount of incorrect addition to the N terminus on one end. We show that because different combinations of stagger and quaternary structure affect the structural symmetry of the fibril end, we propose that differences in quaternary structures will affect directional growth patterns and possibly different morphologies in the mature fiber.
Figures
References
-
- Dobson CM. Protein folding and misfolding. Nature. 2003;426:884–890. - PubMed
-
- Dobson CM. Principles of protein folding, misfolding and aggregation. Semin. Cell Dev. Biol. 2004;15:3–16. - PubMed
-
- Bucciantini M, Giannoni E, Chiti F, Baroni F, Formigli L, Zurdo J, et al. Inherent toxicity of aggregates implies a common mechanism for protein misfolding diseases. Nature. 2002;416:507–511. - PubMed
-
- Stefani M, Dobson CM. Protein aggregation and aggregate toxicity: new insights into protein folding, misfolding diseases and biological evolution. J. Mol. Med. 2003;81:678–699. - PubMed
-
- Cohen FE, Kelly JW. Therapeutic approaches to protein-misfolding diseases. Nature. 2003;426:905–909. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
