

Calcium-dependent regulation of NEMO nuclear export in response to genotoxic stimuli
- PMID: 17074802
- PMCID: PMC1800799
- DOI: 10.1128/MCB.01772-06
Calcium-dependent regulation of NEMO nuclear export in response to genotoxic stimuli
Abstract
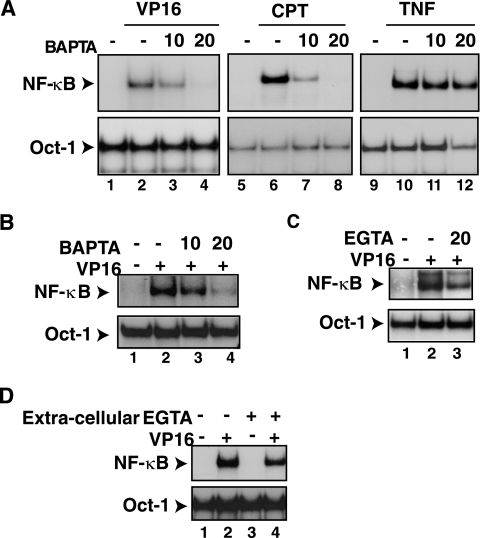
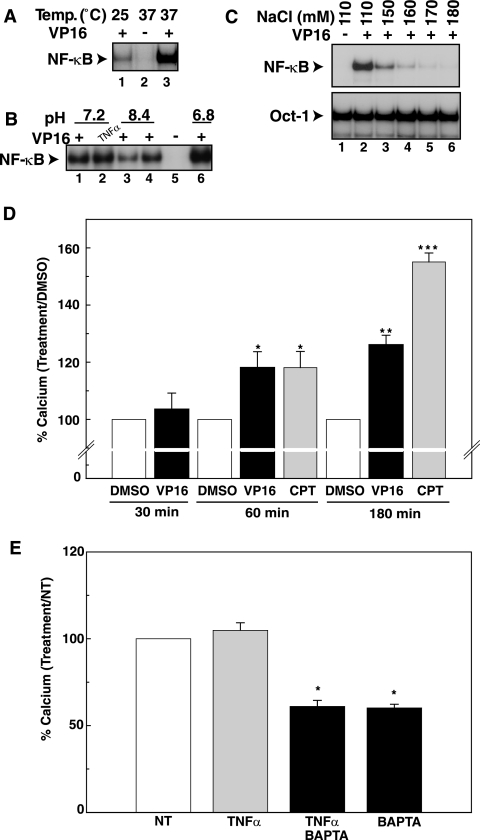
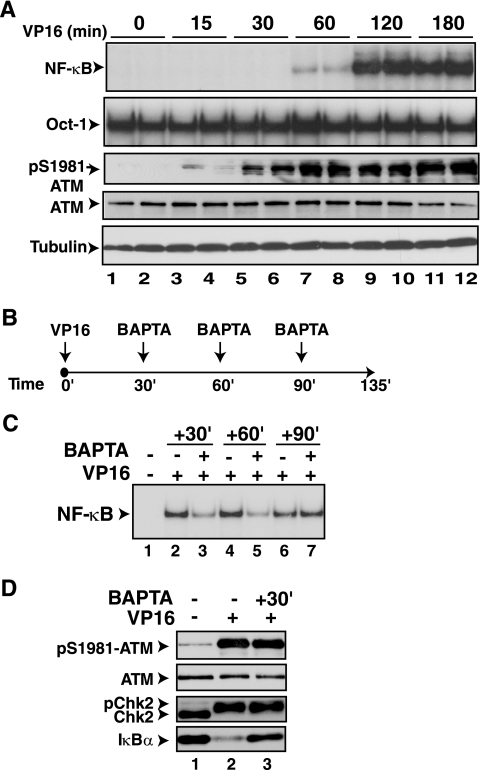
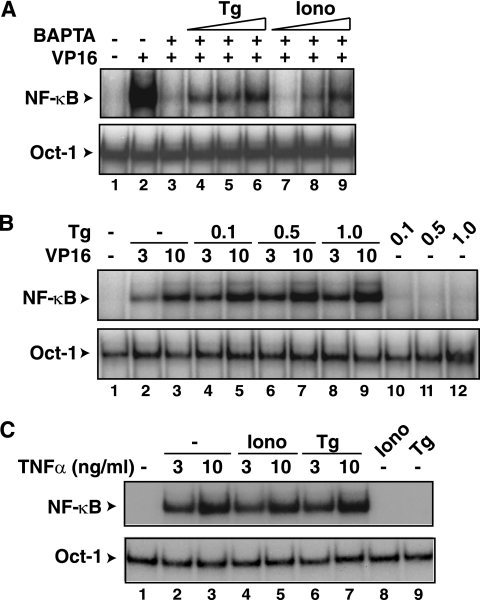
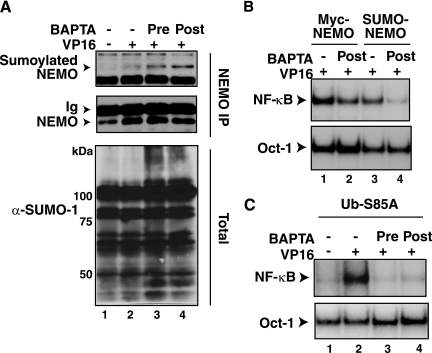
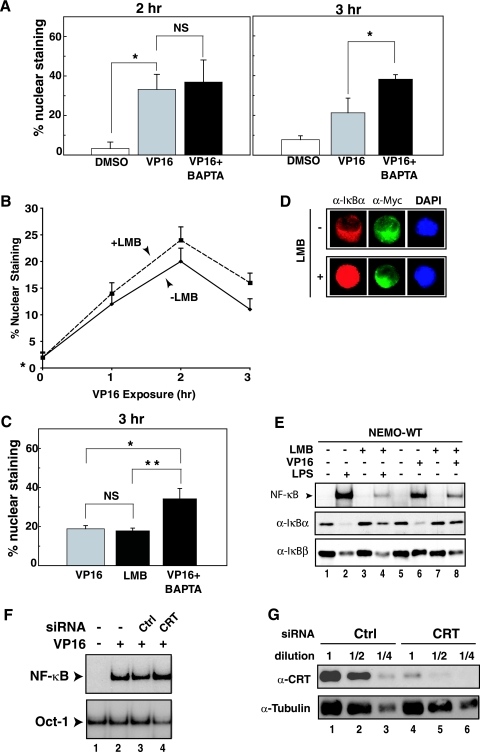
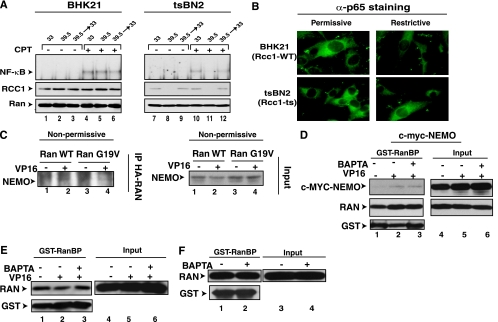
The mechanisms involved in activation of the transcription factor NF-kappaB by genotoxic agents are not well understood. Previously, we provided evidence that a regulatory subunit of the IkappaB kinase (IKK) complex, NF-kappaB essential modulator (NEMO)/IKKgamma, is a component of a nuclear signal that is generated after DNA damage to mediate NF-kappaB activation. Here, we found that etoposide (VP16) and camptothecin induced increases in intracellular free calcium levels at 60 min after stimulation of CEM T leukemic cells. Inhibition of calcium increases by calcium chelators, BAPTA-AM and EGTA-AM, abrogated NF-kappaB activation by these agents in several cell types examined. Conversely, thapsigargin and ionomycin attenuated the BAPTA-AM effects and promoted NF-kappaB activation by the genotoxic stimuli. Analyses of nuclear NEMO levels in VP16-treated cells suggested that calcium was required for nuclear export of NEMO. Inhibition of the nuclear exporter CRM1 by leptomycin B did not interfere with NEMO nuclear export. Similarly, deficiency of a plausible calcium-dependent nuclear export receptor, calreticulin, failed to prevent NF-kappaB activation by VP16. However, temperature inactivation of the Ran guanine nucleotide exchange factor RCC1 in the tsBN2 cell line harboring a temperature-sensitive mutant of RCC1 blocked NF-kappaB activation induced by genotoxic stimuli. Overexpression of Ran in this cell model showed that DNA damage stimuli induced formation of a complex between Ran and NEMO, suggesting that RCC1 regulated NF-kappaB activation through the modulation of RanGTP. Indeed, evidence for VP16-inducible interaction between Ran-GTP and NEMO could be obtained by means of glutathione S-transferase (GST) pull-down assays using GST fused to the Ran binding domain of RanBP2, which specifically interacts with the GTP-bound form of Ran. BAPTA-AM did not alter these interactions, suggesting that calcium is a necessary step beyond the formation of a Ran-GTP-NEMO complex in the nucleus. These results suggest that calcium has a unique role in genotoxic stress-induced NF-kappaB signaling by regulating nuclear export of NEMO subsequent to the formation of a nuclear export complex composed of Ran-GTP, NEMO, and presumably, an undefined nuclear export receptor.
Figures
References
-
- Aggarwal, B. B. 2003. Signalling pathways of the TNFα superfamily: a double-edged sword. Nat. Rev. Immunol. 3:745-756. - PubMed
-
- Akira, S., and K. Takeda. 2004. Toll-like receptor signalling. Nat. Rev. Immunol. 4:499-511. - PubMed
-
- Amundson, S. A., M. Bittner, and A. J. Fornace. 2003. Functional genomics as a window on radiation stress signaling. Oncogene 22:5828-5833. - PubMed
-
- Bakkenist, C. J., and M. B. Kastan. 2003. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature 421:499-506. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous