

Ionic mechanisms of cardiac cell swelling induced by blocking Na+/K+ pump as revealed by experiments and simulation
- PMID: 17074975
- PMCID: PMC2151580
- DOI: 10.1085/jgp.200609646
Ionic mechanisms of cardiac cell swelling induced by blocking Na+/K+ pump as revealed by experiments and simulation
Abstract
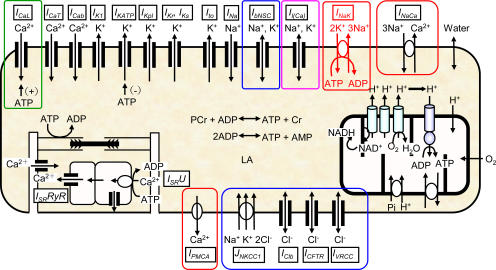
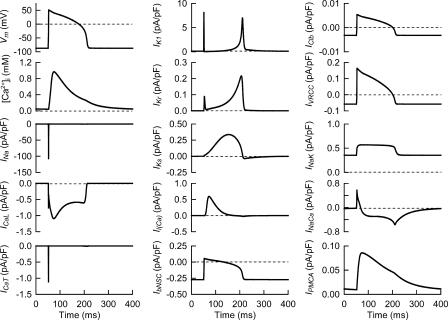
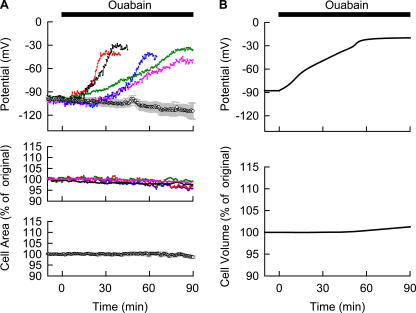
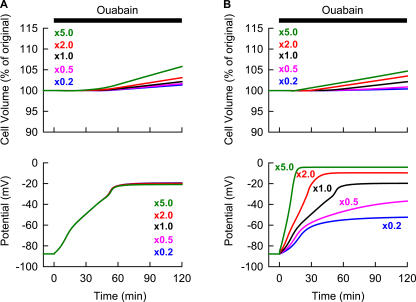
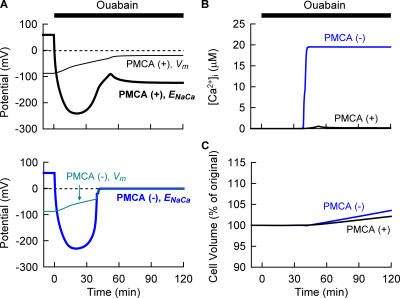
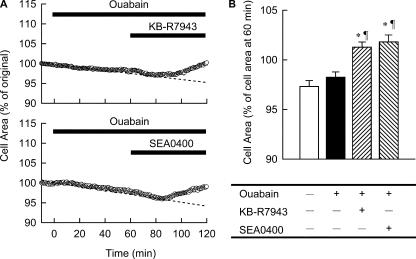
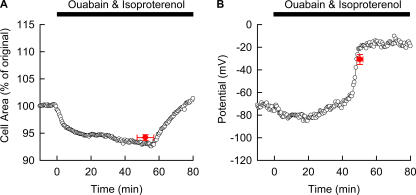
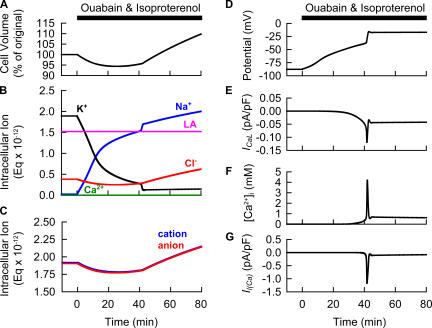
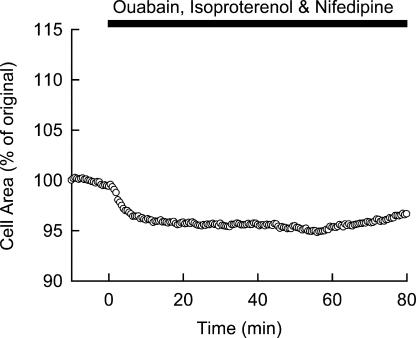
Although the Na(+)/K(+) pump is one of the key mechanisms responsible for maintaining cell volume, we have observed experimentally that cell volume remained almost constant during 90 min exposure of guinea pig ventricular myocytes to ouabain. Simulation of this finding using a comprehensive cardiac cell model (Kyoto model incorporating Cl(-) and water fluxes) predicted roles for the plasma membrane Ca(2+)-ATPase (PMCA) and Na(+)/Ca(2+) exchanger, in addition to low membrane permeabilities for Na(+) and Cl(-), in maintaining cell volume. PMCA might help maintain the [Ca(2+)] gradient across the membrane though compromised, and thereby promote reverse Na(+)/Ca(2+) exchange stimulated by the increased [Na(+)](i) as well as the membrane depolarization. Na(+) extrusion via Na(+)/Ca(2+) exchange delayed cell swelling during Na(+)/K(+) pump block. Supporting these model predictions, we observed ventricular cell swelling after blocking Na(+)/Ca(2+) exchange with KB-R7943 or SEA0400 in the presence of ouabain. When Cl(-) conductance via the cystic fibrosis transmembrane conductance regulator (CFTR) was activated with isoproterenol during the ouabain treatment, cells showed an initial shrinkage to 94.2 +/- 0.5%, followed by a marked swelling 52.0 +/- 4.9 min after drug application. Concomitantly with the onset of swelling, a rapid jump of membrane potential was observed. These experimental observations could be reproduced well by the model simulations. Namely, the Cl(-) efflux via CFTR accompanied by a concomitant cation efflux caused the initial volume decrease. Then, the gradual membrane depolarization induced by the Na(+)/K(+) pump block activated the window current of the L-type Ca(2+) current, which increased [Ca(2+)](i). Finally, the activation of Ca(2+)-dependent cation conductance induced the jump of membrane potential, and the rapid accumulation of intracellular Na(+) accompanied by the Cl(-) influx via CFTR, resulting in the cell swelling. The pivotal role of L-type Ca(2+) channels predicted in the simulation was demonstrated in experiments, where blocking Ca(2+) channels resulted in a much delayed cell swelling.
Figures
Comment in
-
Cell volume regulation in cardiac myocytes: a leaky boat gets a new bilge pump.J Gen Physiol. 2006 Nov;128(5):487-9. doi: 10.1085/jgp.200609679. J Gen Physiol. 2006. PMID: 17074973 Free PMC article. No abstract available.
References
-
- Akar, J.G., T.H. Everett, R. Ho, J.C. Craft, D.E. Haines, A.P. Somlyo, and A.V. Somlyo. 2003. Intracellular chloride accumulation and subcellular elemental distribution during atrial fibrillation. Circulation. 107:1810–1815. - PubMed
-
- Bahinski, A., A.C. Nairn, P. Greengard, and D.C. Gadsby. 1989. Chloride conductance regulated by cyclic AMP-dependent protein kinase in cardiac myocytes. Nature. 340:718–721. - PubMed
-
- Bak, M.I., and J.S. Ingwall. 2003. Contribution of Na+/H+ exchange to Na+ overload in the ischemic hypertrophied hyperthyroid rat heart. Cardiovasc. Res. 57:1004–1014. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
