

Inefficient human immunodeficiency virus replication in mobile lymphocytes
- PMID: 17079292
- PMCID: PMC1797449
- DOI: 10.1128/JVI.01629-06
Inefficient human immunodeficiency virus replication in mobile lymphocytes
Abstract
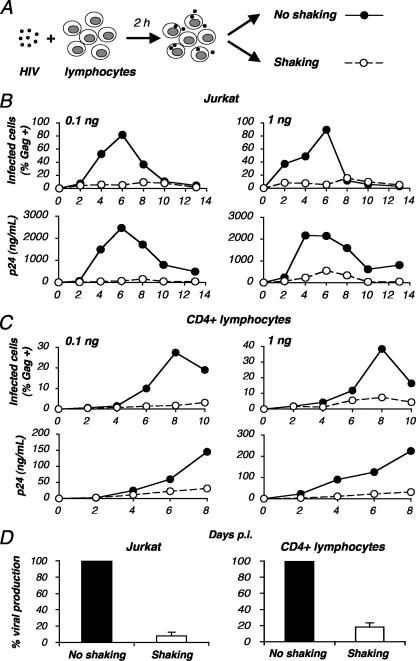
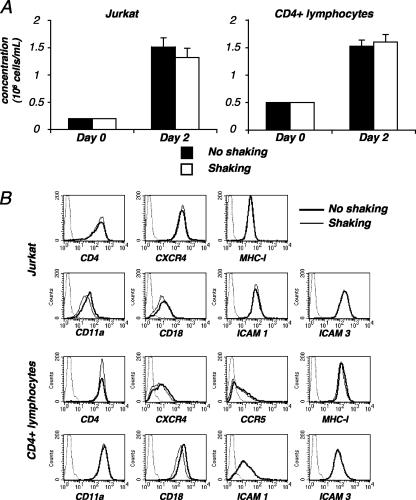
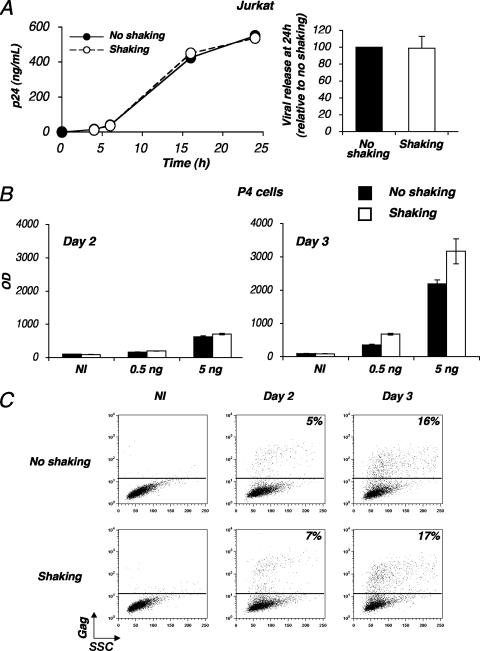
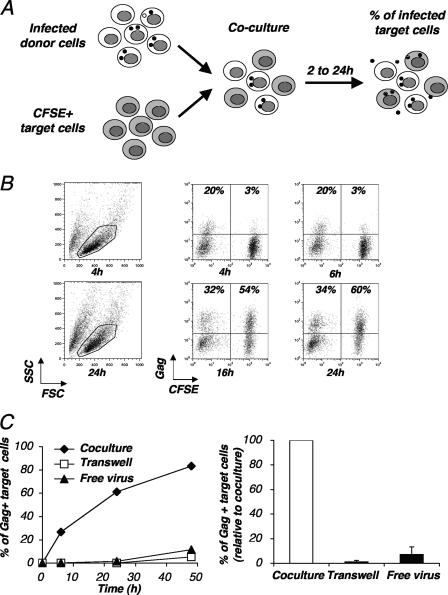
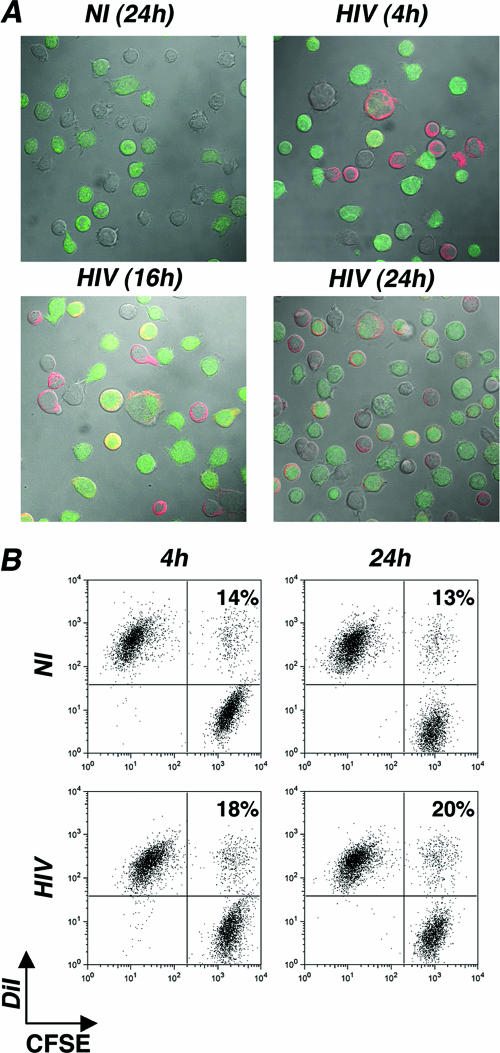
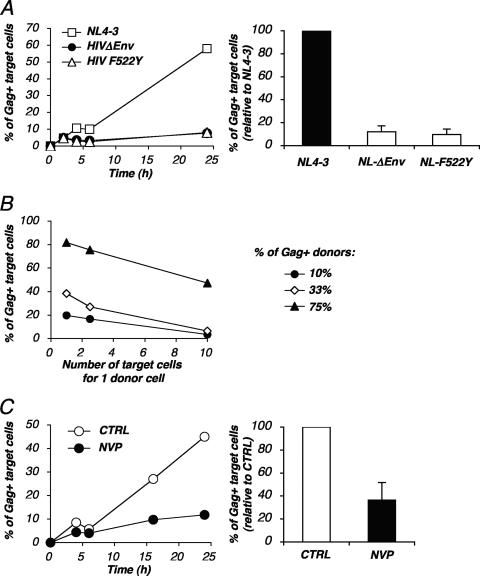
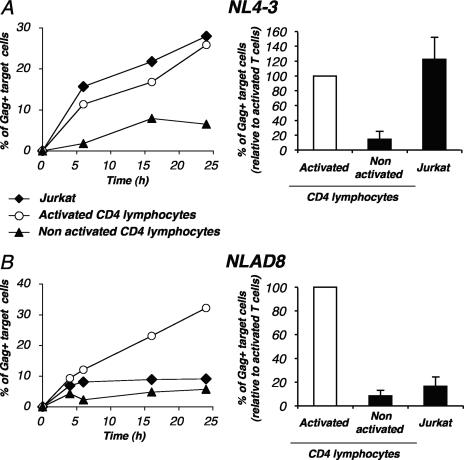
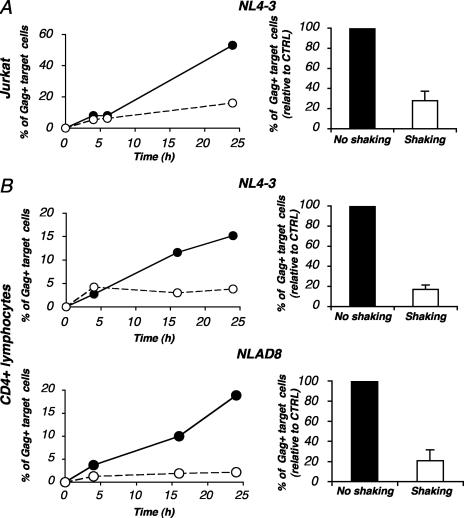
Cell-to-cell viral transfer facilitates the spread of lymphotropic retroviruses such as human immunodeficiency virus (HIV) and human T-cell leukemia virus (HTLV), likely through the formation of "virological synapses" between donor and target cells. Regarding HIV replication, the importance of cell contacts has been demonstrated, but this phenomenon remains only partly characterized. In order to alter cell-to-cell HIV transmission, we have maintained cultures under continuous gentle shaking and followed viral replication in this experimental system. In lymphoid cell lines, as well as in primary lymphocytes, viral replication was dramatically reduced in shaken cultures. To document this phenomenon, we have developed an assay to assess the relative contributions of free and cell-associated virions in HIV propagation. Acutely infected donor cells were mixed with carboxyfluorescein diacetate succinimidyl ester-labeled lymphocytes as targets, and viral production was followed by measuring HIV Gag expression at different time points by flow cytometry. We report that cellular contacts drastically enhance productive viral transfer compared to what is seen with infection with free virus. Productive cell-to-cell viral transmission required fusogenic viral envelope glycoproteins on donor cells and adequate receptors on targets. Only a few syncytia were observed in this coculture system. Virus release from donor cells was unaffected when cultures were gently shaken, whereas virus transfer to recipient cells was severely impaired. Altogether, these results indicate that cell-to-cell transfer is the predominant mode of HIV spread and help to explain why this virus replicates so efficiently in lymphoid organs.
Figures
Similar articles
-
Preliminary in vitro growth cycle and transmission studies of HIV-1 in an autologous primary cell assay of blood-derived macrophages and peripheral blood mononuclear cells.Virology. 1996 Dec 15;226(2):205-16. doi: 10.1006/viro.1996.0648. Virology. 1996. PMID: 8955040
-
A clathrin-dynamin-dependent endocytic pathway for the uptake of HIV-1 by direct T cell-T cell transmission.Antiviral Res. 2008 Nov;80(2):185-93. doi: 10.1016/j.antiviral.2008.06.004. Epub 2008 Jul 3. Antiviral Res. 2008. PMID: 18602423
-
Nuclear localization of Vpr is crucial for the efficient replication of HIV-1 in primary CD4+ T cells.Virology. 2004 Oct 1;327(2):249-61. doi: 10.1016/j.virol.2004.06.024. Virology. 2004. PMID: 15351213
-
[Chemokine receptors and its importance in the replication cycle of human immunodeficiency virus: clinical and therapeutic implications].Acta Med Port. 2008 Sep-Oct;21(5):497-504. Epub 2009 Jan 16. Acta Med Port. 2008. PMID: 19187693 Review. Portuguese.
-
HIV-1 at the immunological and T-lymphocytic virological synapse.Biol Chem. 2008 Oct;389(10):1253-60. doi: 10.1515/BC.2008.143. Biol Chem. 2008. PMID: 18713012 Review.
Cited by
-
The immunological synapse: the gateway to the HIV reservoir.Immunol Rev. 2013 Jul;254(1):305-25. doi: 10.1111/imr.12080. Immunol Rev. 2013. PMID: 23772628 Free PMC article. Review.
-
Immunological Paradigms, Mechanisms, and Models: Conceptual Understanding Is a Prerequisite to Effective Modeling.Front Immunol. 2019 Nov 5;10:2522. doi: 10.3389/fimmu.2019.02522. eCollection 2019. Front Immunol. 2019. PMID: 31749803 Free PMC article. Review.
-
Human immunodeficiency virus type 1 and influenza virus exit via different membrane microdomains.J Virol. 2007 Nov;81(22):12630-40. doi: 10.1128/JVI.01255-07. Epub 2007 Sep 12. J Virol. 2007. PMID: 17855546 Free PMC article.
-
Restriction of HIV-1 replication in macrophages and CD4+ T cells from HIV controllers.Blood. 2011 Jul 28;118(4):955-64. doi: 10.1182/blood-2010-12-327106. Epub 2011 Jun 3. Blood. 2011. PMID: 21642597 Free PMC article.
-
Cell-to-cell transmission of retroviruses: Innate immunity and interferon-induced restriction factors.Virology. 2011 Mar 15;411(2):251-9. doi: 10.1016/j.virol.2010.12.031. Epub 2011 Jan 17. Virology. 2011. PMID: 21247613 Free PMC article. Review.
References
-
- Barnard, A. L., T. Igakura, Y. Tanaka, G. P. Taylor, and C. R. Bangham. 2005. Engagement of specific T-cell surface molecules regulates cytoskeletal polarization in HTLV-1-infected lymphocytes. Blood 106:988-995. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
