

The varicella-zoster virus (VZV) ORF9 protein interacts with the IE62 major VZV transactivator
- PMID: 17079304
- PMCID: PMC1797441
- DOI: 10.1128/JVI.01274-06
The varicella-zoster virus (VZV) ORF9 protein interacts with the IE62 major VZV transactivator
Abstract
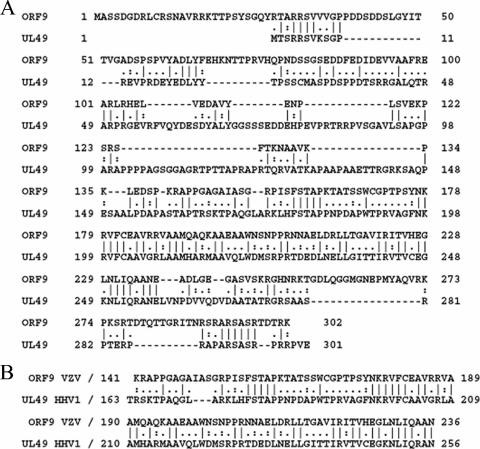
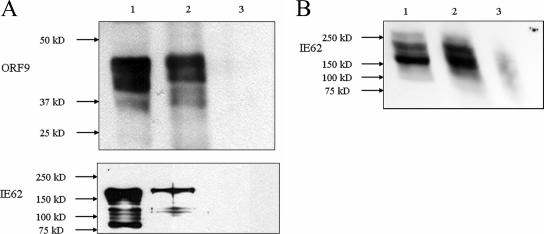
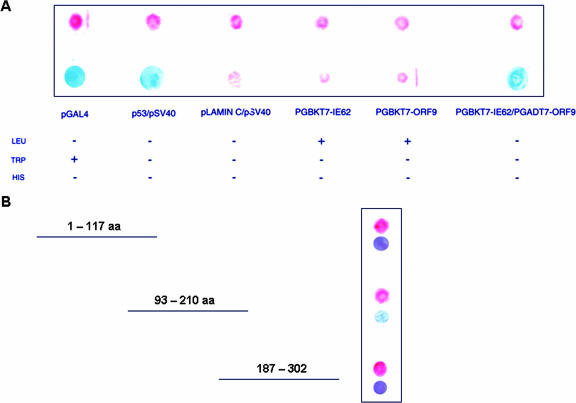
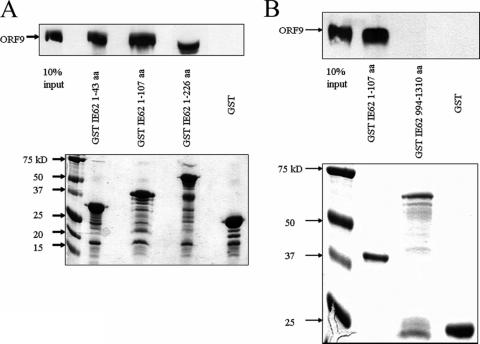
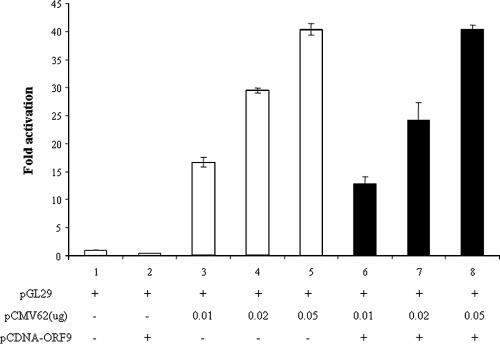
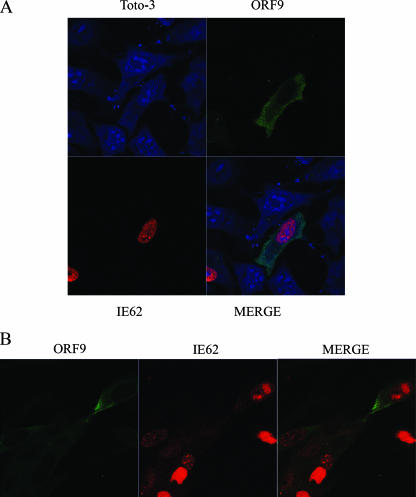
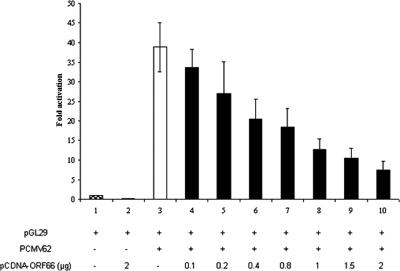
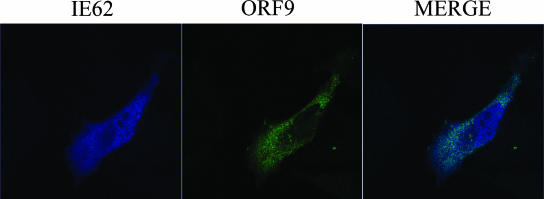
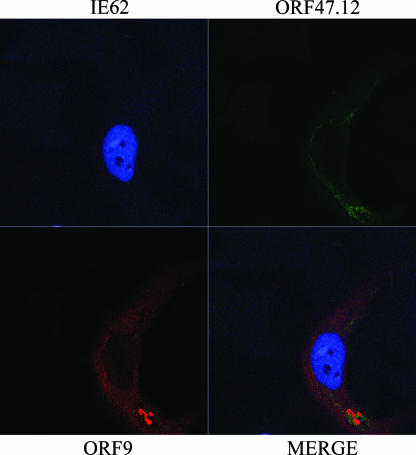
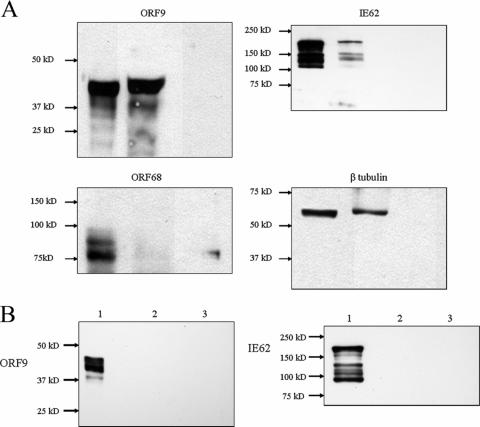
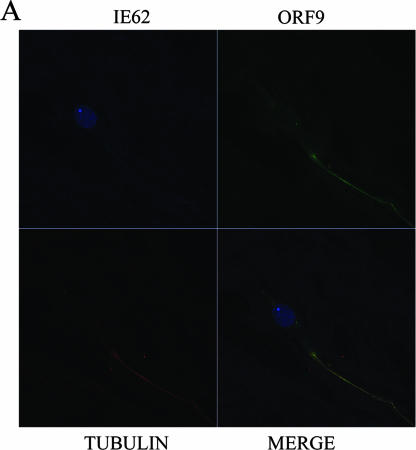
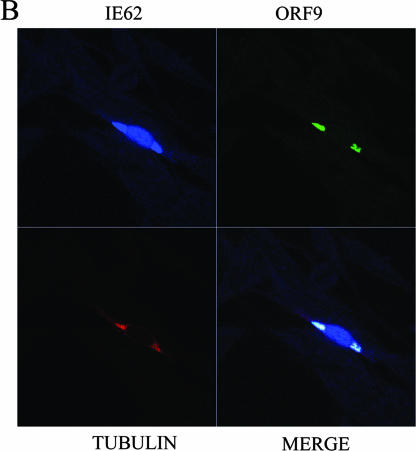
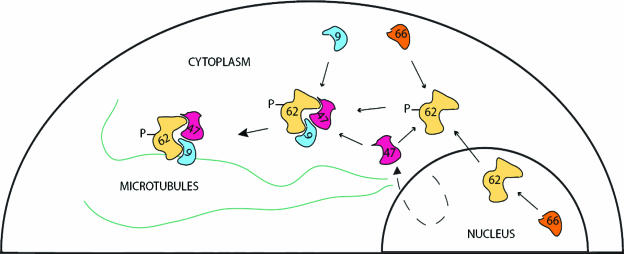
The varicella-zoster virus (VZV) ORF9 protein is a member of the herpesvirus UL49 gene family but shares limited identity and similarity with the UL49 prototype, herpes simplex virus type 1 VP22. ORF9 mRNA is the most abundantly expressed message during VZV infection; however, little is known concerning the functions of the ORF9 protein. We have found that the VZV major transactivator IE62 and the ORF9 protein can be coprecipitated from infected cells. Yeast two-hybrid analysis localized the region of the ORF9 protein required for interaction with IE62 to the middle third of the protein encompassing amino acids 117 to 186. Protein pull-down assays with GST-IE62 fusion proteins containing N-terminal IE62 sequences showed that amino acids 1 to 43 of the acidic transcriptional activation domain of IE62 can bind recombinant ORF9 protein. Confocal microscopy of transiently transfected cells showed that in the absence of other viral proteins, the ORF9 protein was localized in the cytoplasm while IE62 was localized in the nucleus. In VZV-infected cells, the ORF9 protein was localized to the cytoplasm whereas IE62 exhibited both nuclear and cytoplasmic localization. Cotransfection of plasmids expressing ORF9, IE62, and the viral ORF66 kinase resulted in significant colocalization of ORF9 and IE62 in the cytoplasm. Coimmunoprecipitation experiments with antitubulin antibodies indicate the presence of ORF9-IE62-tubulin complexes in infected cells. Colocalization of ORF9 and tubulin in transfected cells was visualized by confocal microscopy. These data suggest a model for ORF9 protein function involving complex formation with IE62 and possibly other tegument proteins in the cytoplasm at late times in infection.
Figures
Similar articles
-
Phosphorylation of the varicella-zoster virus (VZV) major transcriptional regulatory protein IE62 by the VZV open reading frame 66 protein kinase.J Virol. 2006 Feb;80(4):1710-23. doi: 10.1128/JVI.80.4.1710-1723.2006. J Virol. 2006. PMID: 16439528 Free PMC article.
-
Nuclear accumulation of IE62, the varicella-zoster virus (VZV) major transcriptional regulatory protein, is inhibited by phosphorylation mediated by the VZV open reading frame 66 protein kinase.J Virol. 2000 Mar;74(5):2265-77. doi: 10.1128/jvi.74.5.2265-2277.2000. J Virol. 2000. PMID: 10666257 Free PMC article.
-
Intracellular distribution of the ORF4 gene product of varicella-zoster virus is influenced by the IE62 protein.J Gen Virol. 1996 Jul;77 ( Pt 7):1505-13. doi: 10.1099/0022-1317-77-7-1505. J Gen Virol. 1996. PMID: 8757993
-
Varicella-zoster virus open reading frame 66 protein kinase and its relationship to alphaherpesvirus US3 kinases.Curr Top Microbiol Immunol. 2010;342:79-98. doi: 10.1007/82_2009_7. Curr Top Microbiol Immunol. 2010. PMID: 20186610 Free PMC article. Review.
-
VZV ORF47 serine protein kinase and its viral substrates.Curr Top Microbiol Immunol. 2010;342:99-111. doi: 10.1007/82_2009_5. Curr Top Microbiol Immunol. 2010. PMID: 20186612 Review.
Cited by
-
Sex differences underlying orofacial varicella zoster associated pain in rats.BMC Neurol. 2017 May 17;17(1):95. doi: 10.1186/s12883-017-0882-6. BMC Neurol. 2017. PMID: 28514943 Free PMC article.
-
Simian varicella virus: molecular virology.Curr Top Microbiol Immunol. 2010;342:291-308. doi: 10.1007/82_2010_27. Curr Top Microbiol Immunol. 2010. PMID: 20369316 Free PMC article. Review.
-
Analysis of IE62 mutations found in Varicella-Zoster virus vaccine strains for transactivation activity.J Microbiol. 2018 Jun;56(6):441-448. doi: 10.1007/s12275-018-8144-x. Epub 2018 Jun 1. J Microbiol. 2018. PMID: 29858833
-
Effector and Central Memory Poly-Functional CD4(+) and CD8(+) T Cells are Boosted upon ZOSTAVAX(®) Vaccination.Front Immunol. 2015 Oct 29;6:553. doi: 10.3389/fimmu.2015.00553. eCollection 2015. Front Immunol. 2015. PMID: 26579128 Free PMC article.
-
Varicella-Zoster Virus ORF9p Binding to Cellular Adaptor Protein Complex 1 Is Important for Viral Infectivity.J Virol. 2018 Jul 17;92(15):e00295-18. doi: 10.1128/JVI.00295-18. Print 2018 Aug 1. J Virol. 2018. PMID: 29793951 Free PMC article.
References
-
- Aints, A., H. Guven, G. Gahrton, C. I. Smith, and M. S. Dilber. 2001. Mapping of herpes simplex virus-1 VP22 functional domains for inter- and subcellular protein targeting. Gene Ther. 8:1051-1056. - PubMed
-
- Baiker, A., C. Bagowski, H. Ito, M. Sommer, L. Zerboni, K. Fabel, J. Hay, W. Ruyechan, and A. M. Arvin. 2004. The immediate-early 63 protein of varicella-zoster virus: analysis of functional domains required for replication in vitro and for T-cell and skin tropism in the SCIDhu model in vivo. J. Virol. 78:1181-1194. - PMC - PubMed
-
- Besser, J., M. H. Sommer, L. Zerboni, C. P. Bagowski, H. Ito, J. Moffat, C. C. Ku, and A. M. Arvin. 2003. Differentiation of varicella-zoster virus ORF47 protein kinase and IE62 protein binding domains and their contributions to replication in human skin xenografts in the SCID-hu mouse. J. Virol. 77:5964-5974. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
