

Mutation of beta-glucosidase 2 causes glycolipid storage disease and impaired male fertility
- PMID: 17080196
- PMCID: PMC1626112
- DOI: 10.1172/JCI29224
Mutation of beta-glucosidase 2 causes glycolipid storage disease and impaired male fertility
Abstract
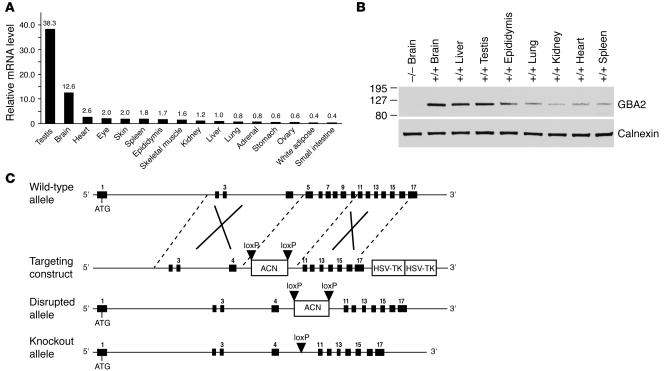
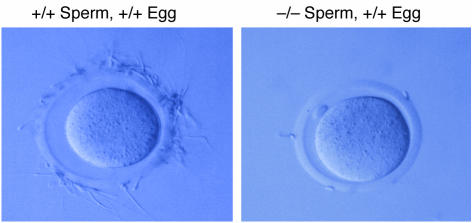
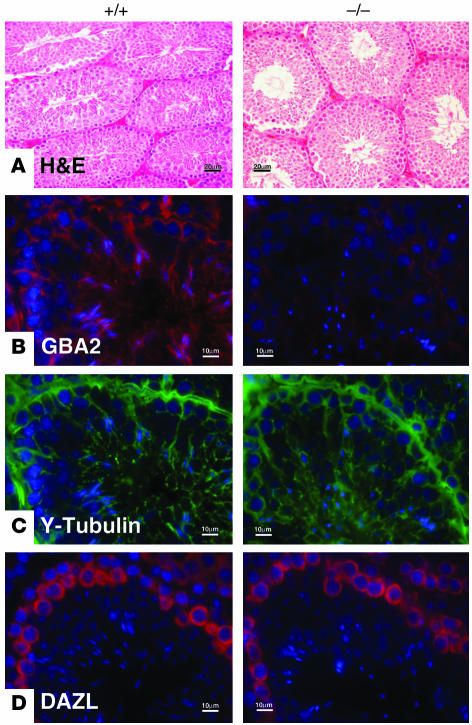
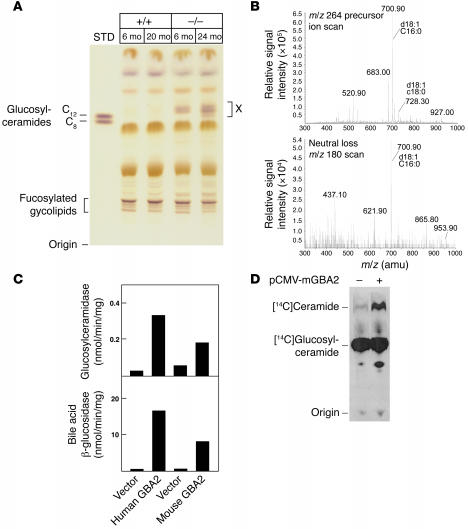
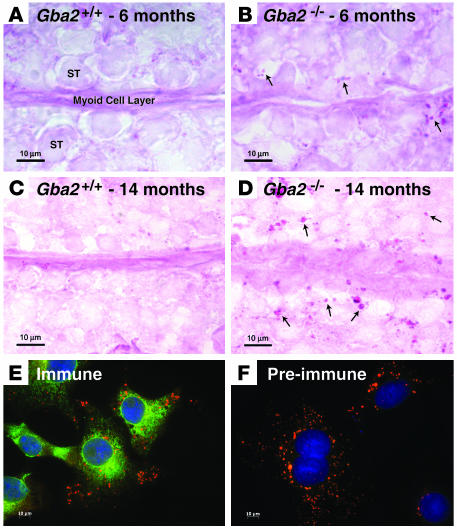
beta-Glucosidase 2 (GBA2) is a resident enzyme of the endoplasmic reticulum thought to play a role in the metabolism of bile acid-glucose conjugates. To gain insight into the biological function of this enzyme and its substrates, we generated mice deficient in GBA2 and found that these animals had normal bile acid metabolism. Knockout males exhibited impaired fertility. Microscopic examination of sperm revealed large round heads (globozoospermia), abnormal acrosomes, and defective mobility. Glycolipids, identified as glucosylceramides by mass spectrometry, accumulated in the testes, brains, and livers of the knockout mice but did not cause obvious neurological symptoms, organomegaly, or a reduction in lifespan. Recombinant GBA2 hydrolyzed glucosylceramide to glucose and ceramide; the same reaction catalyzed by the beta-glucosidase acid 1 (GBA1) defective in subjects with the Gaucher's form of lysosomal storage disease. We conclude that GBA2 is a glucosylceramidase whose loss causes accumulation of glycolipids and an endoplasmic reticulum storage disease.
Figures


Comment in
-
Shaping the sperm head: an ER enzyme leaves its mark.J Clin Invest. 2006 Nov;116(11):2860-3. doi: 10.1172/JCI30221. J Clin Invest. 2006. PMID: 17080191 Free PMC article.
References
-
- Simons K., Toomre D. Lipid rafts and signal transduction. Nat. Rev. Mol. Cell Biol. 2000;1:31–39. - PubMed
-
- Anderson R.G., Jacobson K. A role for lipid shells in targeting proteins to caveolae, rafts, and other lipid domains. Science. 2002;296:1821–1825. - PubMed
-
- Menaldino D.S., et al. Sphingoid bases and de novo ceramide synthesis: enzymes involved, pharmacology, and mechanisms of action. Pharmacol. Res. 2003;47:373–381. - PubMed
-
- Merrill J.A.H. De novo sphingolipid biosynthesis: a necessary, but dangerous pathway. J. Biol. Chem. 2002;277:25843–25846. - PubMed
-
- Perry R.J., Ridgway N.D. Molecular mechanisms of ceramide transport. Biochim. Biophys. Acta. 2005;1734:220–234. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
