

Deregulated replication licensing causes DNA fragmentation consistent with head-to-tail fork collision
- PMID: 17081992
- PMCID: PMC1819398
- DOI: 10.1016/j.molcel.2006.09.010
Deregulated replication licensing causes DNA fragmentation consistent with head-to-tail fork collision
Abstract
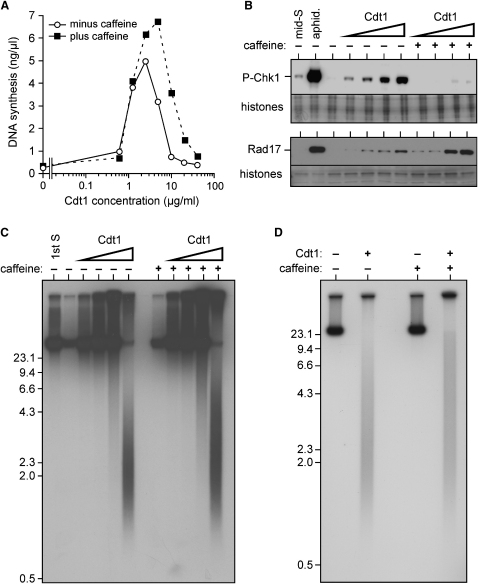
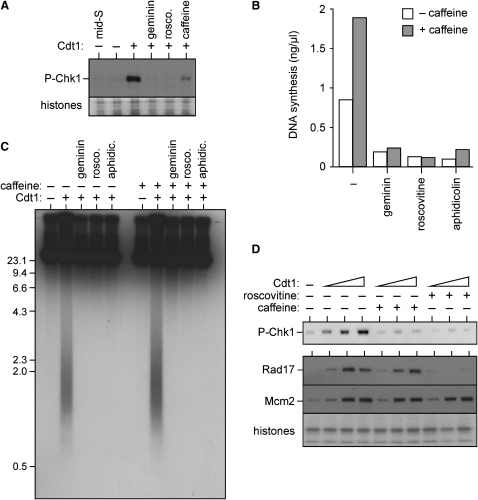
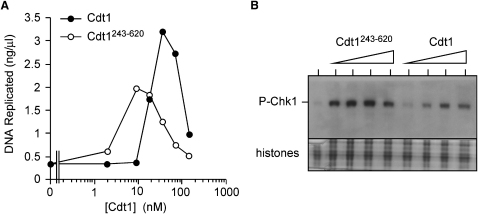
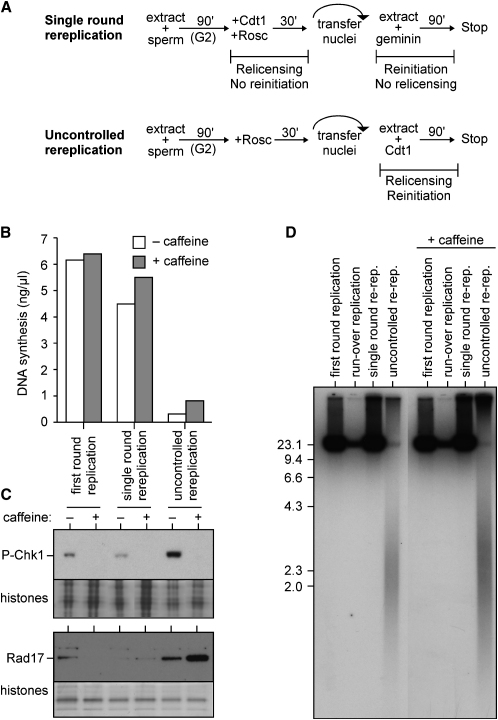
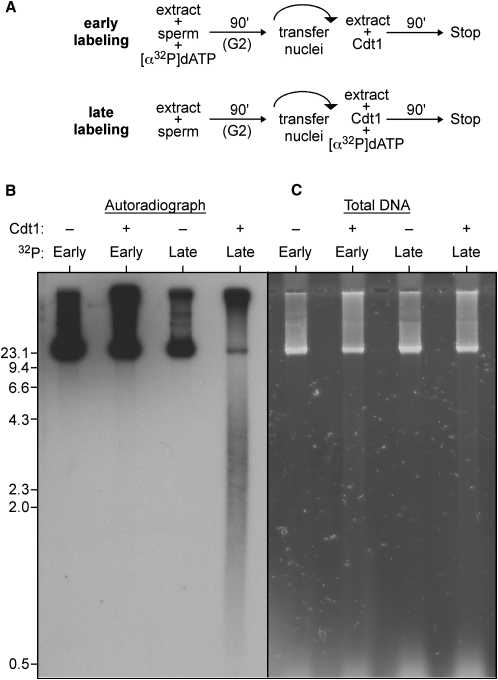
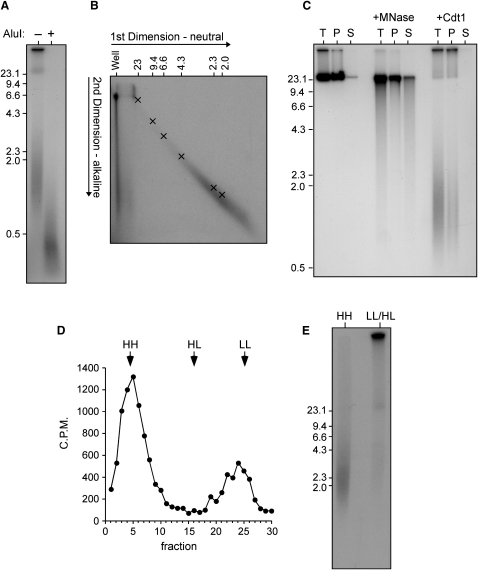
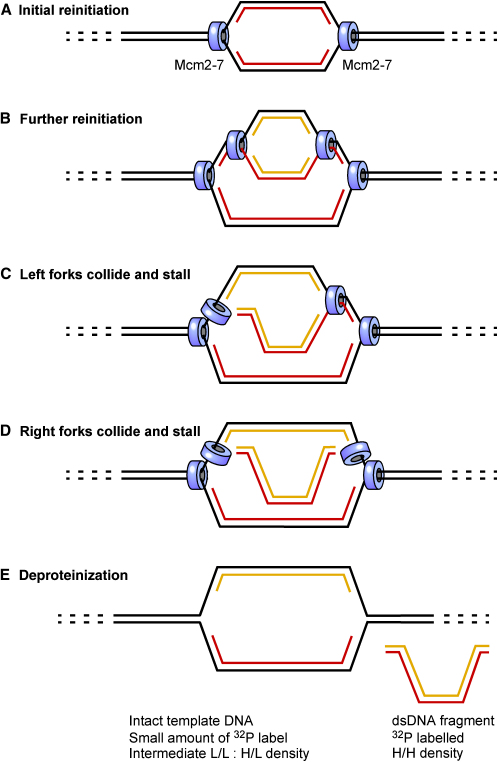
Correct regulation of the replication licensing system ensures that no DNA is rereplicated in a single cell cycle. When the licensing protein Cdt1 is overexpressed in G2 phase of the cell cycle, replication origins are relicensed and the DNA is rereplicated. At the same time, checkpoint pathways are activated that block further cell cycle progression. We have studied the consequence of deregulating the licensing system by adding recombinant Cdt1 to Xenopus egg extracts. We show that Cdt1 induces checkpoint activation and the appearance of small fragments of double-stranded DNA. DNA fragmentation and strong checkpoint activation are dependent on uncontrolled rereplication and do not occur after a single coordinated round of rereplication. The DNA fragments are composed exclusively of rereplicated DNA. The unusual characteristics of these fragments suggest that they result from head-to-tail collision (rear ending) of replication forks chasing one another along the same DNA template.
Figures
References
-
- Annunziato A.T., Seale R.L. Histone deacetylation is required for the maturation of newly replicated chromatin. J. Biol. Chem. 1983;258:12675–12684. - PubMed
-
- Blow J.J., Laskey R.A. A role for the nuclear envelope in controlling DNA replication within the cell cycle. Nature. 1988;332:546–548. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
