

Ferroportin-mediated mobilization of ferritin iron precedes ferritin degradation by the proteasome
- PMID: 17082767
- PMCID: PMC1636618
- DOI: 10.1038/sj.emboj.7601409
Ferroportin-mediated mobilization of ferritin iron precedes ferritin degradation by the proteasome
Abstract
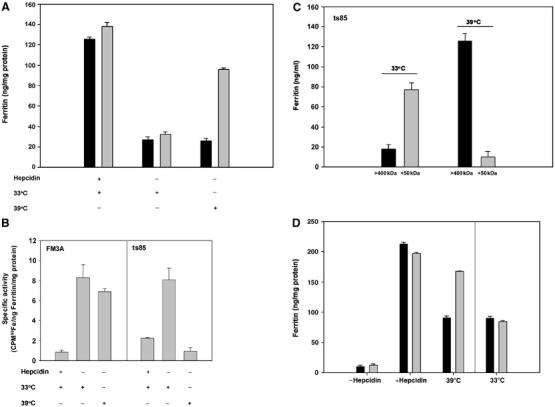
Ferritin is a cytosolic molecule comprised of subunits that self-assemble into a nanocage capable of containing up to 4500 iron atoms. Iron stored within ferritin can be mobilized for use within cells or exported from cells. Expression of ferroportin (Fpn) results in export of cytosolic iron and ferritin degradation. Fpn-mediated iron loss from ferritin occurs in the cytosol and precedes ferritin degradation by the proteasome. Depletion of ferritin iron induces the monoubiquitination of ferritin subunits. Ubiquitination is not required for iron release but is required for disassembly of ferritin nanocages, which is followed by degradation of ferritin by the proteasome. Specific mammalian machinery is not required to extract iron from ferritin. Iron can be removed from ferritin when ferritin is expressed in Saccharomyces cerevisiae, which does not have endogenous ferritin. Expressed ferritin is monoubiquitinated and degraded by the proteasome. Exposure of ubiquitination defective mammalian cells to the iron chelator desferrioxamine leads to degradation of ferritin in the lysosome, which can be prevented by inhibitors of autophagy. Thus, ferritin degradation can occur through two different mechanisms.
Figures




References
-
- Bridges KR (1987) Ascorbic acid inhibits lysosomal autophagy of ferritin. J Biol Chem 262: 14773–14778 - PubMed
-
- Bridges KR, Hoffman KE (1986) The effects of ascorbic acid on the intracellular metabolism of iron and ferritin. J Biol Chem 261: 14273–14277 - PubMed
-
- Crosby WH (1966) Mucosal block. An evaluation of concepts relating to control of iron absorption. Semin Hematol 3: 299–313 - PubMed
-
- Erhardt JG, Estes JE, Pfeiffer CM, Biesalski HK, Craft NE (2004) Combined measurement of ferritin, soluble transferrin receptor, retinol binding protein, and C-reactive protein by an inexpensive, sensitive, and simple sandwich enzyme-linked immunosorbent assay technique. J Nutr 134: 3127–3132 - PubMed
-
- Finley D, Ciechanover A, Varshavsky A (1984) Thermolability of ubiquitin-activating enzyme from the mammalian cell cycle mutant ts85. Cell 37: 43–55 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
