

A molecular characterization of spontaneous frameshift mutagenesis within the trpA gene of Escherichia coli
- PMID: 17084112
- PMCID: PMC1804121
- DOI: 10.1016/j.dnarep.2006.09.007
A molecular characterization of spontaneous frameshift mutagenesis within the trpA gene of Escherichia coli
Abstract
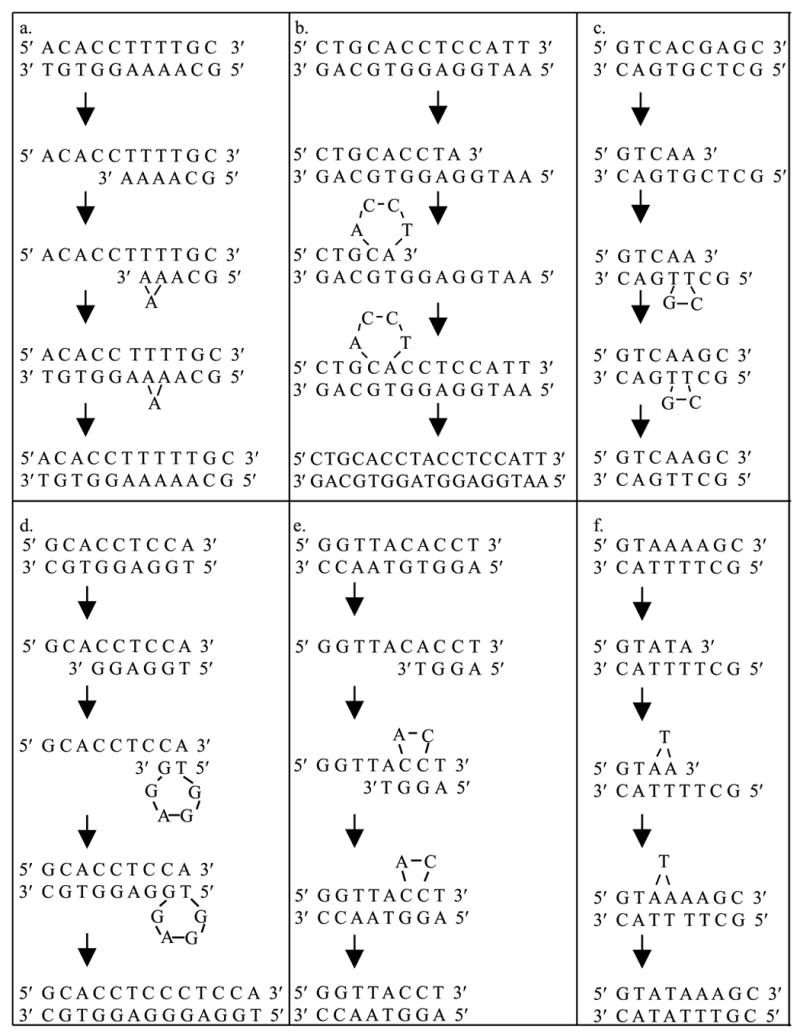
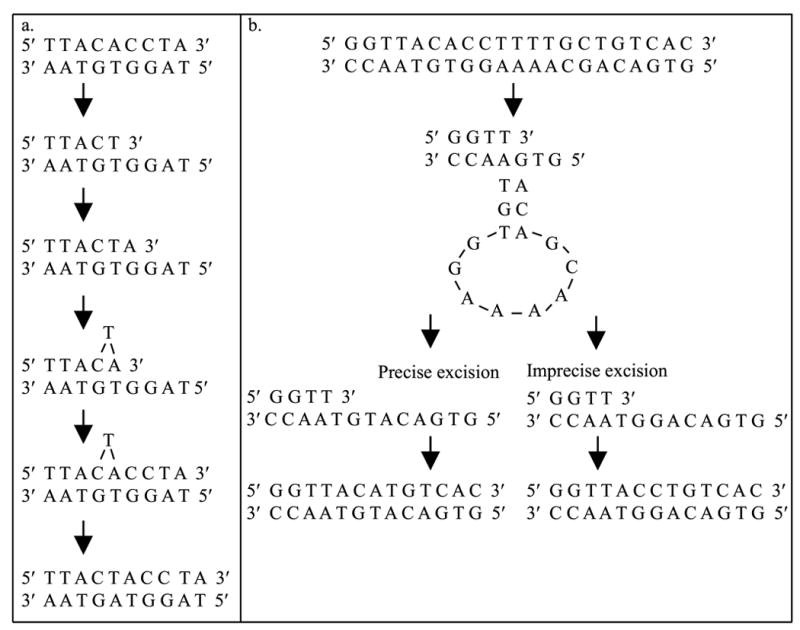
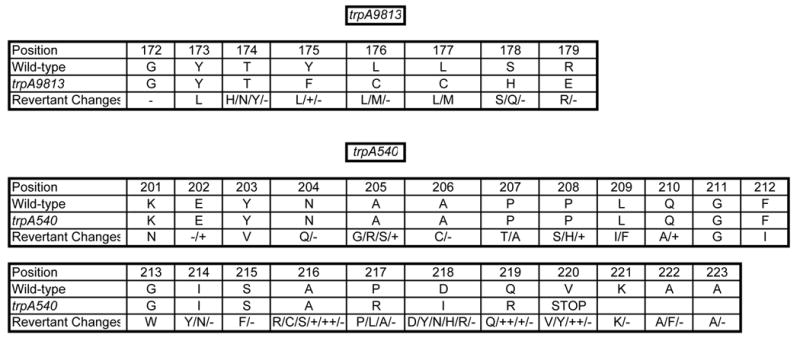
Spontaneous frameshift mutations are an important source of genetic variation in all species and cause a large number of genetic disorders in humans. To enhance our understanding of the molecular mechanisms of frameshift mutagenesis, 583 spontaneous Trp+ revertants of two trpA frameshift alleles in Escherichia coli were isolated and DNA sequenced. In order to measure the contribution of methyl-directed mismatch repair to frameshift production, mutational spectra were constructed for both mismatch repair-proficient and repair-defective strains. The molecular origins of practically all of the frameshifts analyzed could be explained by one of six simple models based upon misalignment of the template or nascent DNA strands with or without misincorporation of primer nucleotides during DNA replication. Most frameshifts occurred within mononucleotide runs as has been shown often in previous studies but the location of the 76 frameshift sites was usually outside of runs. Mismatch repair generally was most effective in preventing the occurrence of frameshifts within runs but there was much variation from site to site. Most frameshift sites outside of runs appear to be refractory to mismatch repair although the small number of occurrences at most of these sites make firm conclusions impossible. There was a dense pattern of reversion sites within the trpA DNA region where reversion events could occur, suggesting that, in general, most DNA sequences are capable of undergoing spontaneous mutational events during replication that can lead to small deletions and insertions. Many of these errors are likely to occur at low frequencies and be tolerated as events too costly to prevent or repair. These studies also revealed an unpredicted flexibility in the primary amino acid sequence of the trpA product, the alpha subunit of tryptophan synthase.
Figures

References
-
- Echols H, Goodman MF. Fidelity mechanisms in DNA replication. Ann Rev Biochem. 1991;60:477–511. - PubMed
-
- Schaaper RM. Base selection, proofreading, and mismatch repair during DNA replication in Escherichia coli. J Biol Chem. 1993;268:23762–23765. - PubMed
-
- Maki H. Origins of spontaneous mutations: specificity and directionality of base substitutions, frameshifts and sequence-substitution mutagenesis. Ann Rev Genet. 2002;36:279–303. - PubMed
-
- Streisinger G, Okada Y, Emrich J, Newton A, Tsugita E, Terzhagi E, Inouye M. Frameshift mutations and the genetic code. Cold Spring Harbor Symp Quant Biol. 1966;31:77–84. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
