

Analysis of two-dimensional dissociation constant of laterally mobile cell adhesion molecules
- PMID: 17085486
- PMCID: PMC1779959
- DOI: 10.1529/biophysj.106.089649
Analysis of two-dimensional dissociation constant of laterally mobile cell adhesion molecules
Abstract
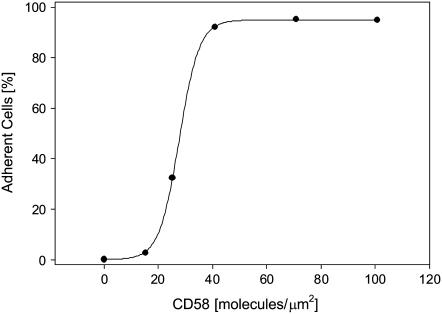
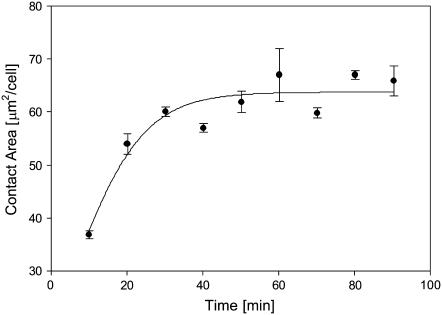
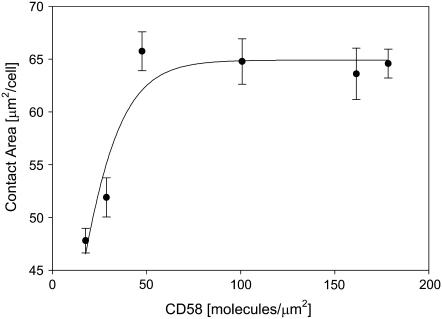
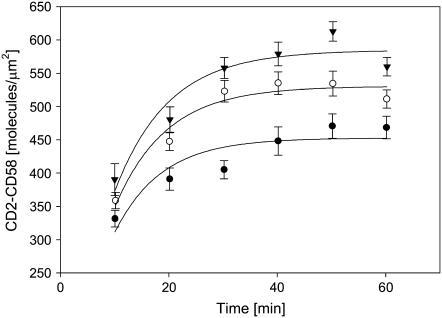
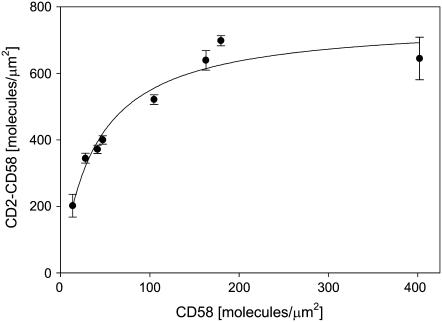
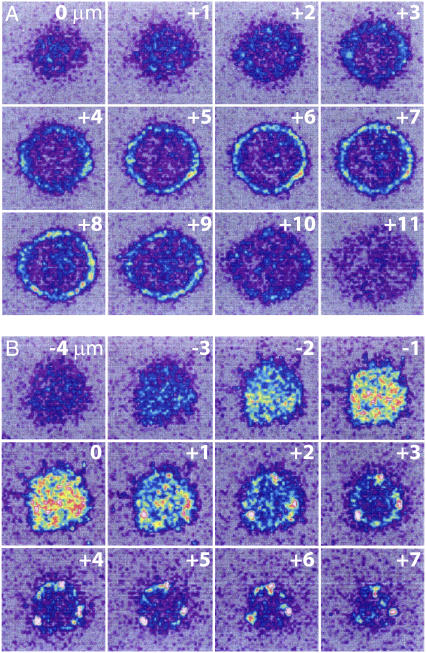
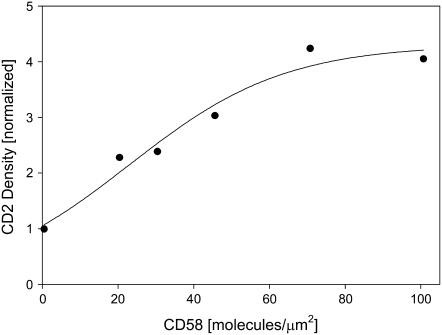
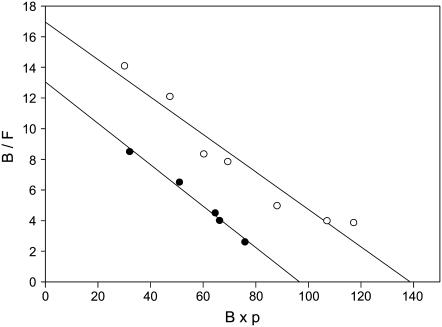
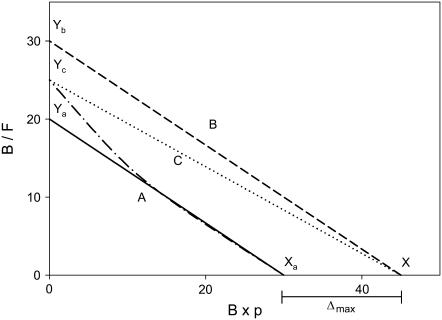
We formulate a general analysis to determine the two-dimensional dissociation constant (2D Kd), and use this method to study the interaction of CD2-expressing T cells with glass-supported planar bilayers containing fluorescently labeled CD58, a CD2 counter-receptor. Both CD2 and CD58 are laterally mobile in their respective membranes. Adhesion is indicated by accumulation of CD2 and CD58 in the cell-bilayer contact area; adhesion molecule density and contact area size attain equilibrium within 40 min. The standard (Scatchard) analysis of solution-phase binding is not applicable to the case of laterally mobile adhesion molecules due to the dynamic nature of the interaction. We derive a new binding equation, B/F=[(Ntxf)/(KdxScell)]-[(Bxp)/Kd], where B and F are bound and free CD58 density in the contact area, respectively; Nt is CD2 molecule number per cell; f is CD2 fractional mobility; Scell is cell surface area; and p is the ratio of contact area at equilibrium to Scell. We use this analysis to determine that the 2D Kd for CD2-CD58 is 5.4-7.6 molecules/microm2. 2D Kd analysis provides a general and quantitative measure of the mechanisms regulating cell-cell adhesion.
Figures
References
-
- Dustin, M. L., S. K. Bromley, M. M. Davis, and C. Zhu. 2001. Identification of self through two-dimensional chemistry and synapses. Annu. Rev. Cell Dev. Biol. 17:133–157. - PubMed
-
- Davis, S. J., S. Ikemizu, M. K. Wild, and P. A. van der Merwe. 1998. CD2 and the nature of protein interactions mediating cell-cell recognition. Immunol. Rev. 163:217–236. - PubMed
-
- Tangye, S. G., J. H. Phillips, and L. L. Lanier. 2000. The CD2-subset of the Ig superfamily of cell surface molecules: receptor-ligand pairs expressed by NK cells and other immune cells. Semin. Immunol. 12:149–157. - PubMed
-
- Doyle, C., and J. L. Strominger. 1987. Interaction between CD4 and class-II MHC molecules mediates cell-adhesion. Nature. 330:256–259. - PubMed
-
- Luescher, I. F., E. Vivier, A. Layer, J. Mahiou, F. Godeau, B. Malissen, and P. Romero. 1995. CD8 modulation of T-cell antigen receptor-ligand interactions on living cytotoxic T-lymphocytes. Nature. 373:353–356. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
