

Self-assembling of peptide/membrane complexes by atomistic molecular dynamics simulations
- PMID: 17085495
- PMCID: PMC1779969
- DOI: 10.1529/biophysj.106.093013
Self-assembling of peptide/membrane complexes by atomistic molecular dynamics simulations
Abstract
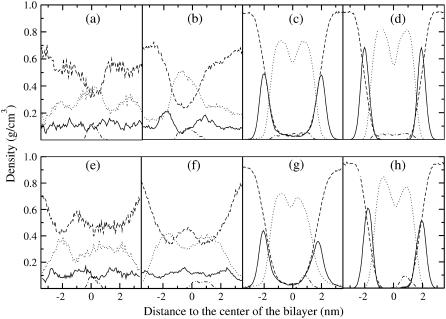
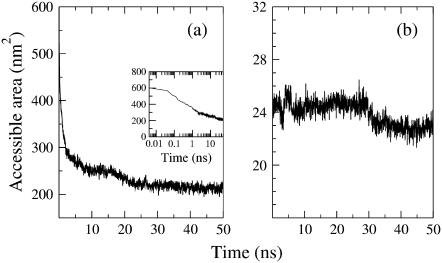
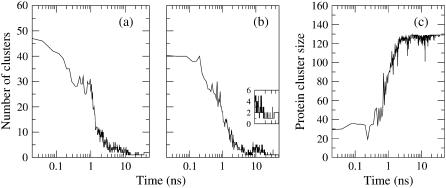
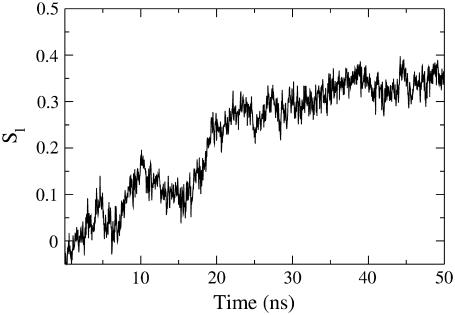
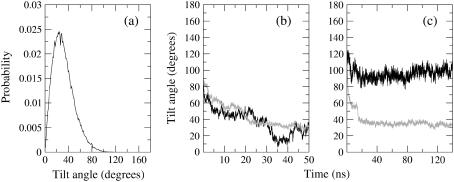
Model biological membranes consisting of peptide/lipid-bilayer complexes can nowadays be studied by classical molecular dynamics (MD) simulations at atomic detail. In most cases, the simulation starts with an assumed state of a peptide in a preformed bilayer, from which equilibrium configurations are difficult to obtain due to a relatively slow molecular diffusion. As an alternative, we propose an extension of reported work on the self-organization of unordered lipids into bilayers, consisting of including a peptide molecule in the initial random configuration to obtain a membrane-bound peptide simultaneous to the formation of the lipid bilayer. This strategy takes advantage of the fast reorganization of lipids, among themselves and around the peptide, in an aqueous environment. Model peptides of different hydrophobicity, CH3-CO-W2L18W2-NH2 (WL22) and CH3-CO-W2A18W2-NH2 (WA22), in dipalmitoyl-phosphatidylcholine (DPPC) are used as test cases. In the equilibrium states of the peptide/membrane complexes, achieved in time ranges of 50-100 ns, the two peptides behave as expected from experimental and theoretical studies. The strongly hydrophobic WL22 is inserted in a transmembrane configuration and the marginally apolar, alanine-based WA22 is found in two alternative states: transmembrane inserted or parallel to the membrane plane, embedded close to the bilayer interface, with similar stability. This shows that the spontaneous assembly of peptides and lipids is an unbiased and reliable strategy to produce and study models of equilibrated peptide/lipid complexes of unknown membrane-binding mode and topology.
Figures

Similar articles
-
Molecular dynamics simulations of model trans-membrane peptides in lipid bilayers: a systematic investigation of hydrophobic mismatch.Biophys J. 2006 Apr 1;90(7):2326-43. doi: 10.1529/biophysj.105.073395. Epub 2006 Jan 20. Biophys J. 2006. PMID: 16428278 Free PMC article.
-
Molecular dynamics simulations of indolicidin association with model lipid bilayers.Biophys J. 2007 Jun 15;92(12):L100-2. doi: 10.1529/biophysj.107.108050. Epub 2007 Apr 6. Biophys J. 2007. PMID: 17416617 Free PMC article.
-
Binding and insertion of alpha-helical anti-microbial peptides in POPC bilayers studied by molecular dynamics simulations.Chem Phys Lipids. 2004 Nov;132(1):113-32. doi: 10.1016/j.chemphyslip.2004.09.011. Chem Phys Lipids. 2004. PMID: 15530453
-
Roles of bilayer material properties in function and distribution of membrane proteins.Annu Rev Biophys Biomol Struct. 2006;35:177-98. doi: 10.1146/annurev.biophys.35.040405.102022. Annu Rev Biophys Biomol Struct. 2006. PMID: 16689633 Review.
-
Modelling of proteins in membranes.Chem Phys Lipids. 2006 Jun;141(1-2):2-29. doi: 10.1016/j.chemphyslip.2006.02.024. Epub 2006 Mar 27. Chem Phys Lipids. 2006. PMID: 16620797 Review.
Cited by
-
Antimicrobial Peptide Mechanism Studied by Scattering-Guided Molecular Dynamics Simulation.J Phys Chem B. 2022 Sep 15;126(36):6922-6935. doi: 10.1021/acs.jpcb.2c03193. Epub 2022 Sep 6. J Phys Chem B. 2022. PMID: 36067064 Free PMC article.
-
Fluorescence spectroscopy and molecular dynamics simulations in studies on the mechanism of membrane destabilization by antimicrobial peptides.Cell Mol Life Sci. 2011 Jul;68(13):2281-301. doi: 10.1007/s00018-011-0719-1. Epub 2011 May 17. Cell Mol Life Sci. 2011. PMID: 21584808 Free PMC article. Review.
-
A lipocentric view of peptide-induced pores.Eur Biophys J. 2011 Apr;40(4):399-415. doi: 10.1007/s00249-011-0693-4. Epub 2011 Mar 26. Eur Biophys J. 2011. PMID: 21442255 Free PMC article. Review.
-
Dynamic Processes and Mechanical Properties of Lipid-Nanoparticle Mixtures.Polymers (Basel). 2023 Apr 9;15(8):1828. doi: 10.3390/polym15081828. Polymers (Basel). 2023. PMID: 37111975 Free PMC article.
-
Antimicrobial and Antibiofilm Activity of a Recombinant Fragment of β-Thymosin of Sea Urchin Paracentrotus lividus.Mar Drugs. 2018 Oct 2;16(10):366. doi: 10.3390/md16100366. Mar Drugs. 2018. PMID: 30279359 Free PMC article.
References
-
- Saiz, L., and M. L. Klein. 2002. Computer simulation studies of model biological membranes. Acc. Chem. Res. 35:482–489. - PubMed
-
- Hansson, T., C. Oostenbrink, and W. F. van Gunsteren. 2002. Molecular dynamics simulations. Curr. Opin. Struct. Biol. 12:190–196. - PubMed
-
- Ash, W. L., M. R. Zlomislic, E. O. Oloo, and D. P. Tieleman. 2004. Computer simulations of membrane proteins. Biochim. Biophys. Acta. 1666:158–189. - PubMed
-
- Nymeyer, H., T. B. Woolf, and A. E. Garcia. 2005. Folding is not required for bilayer insertion: replica exchange simulations of an alpha-helical peptide with an explicit lipid bilayer. Proteins. 59:783–790. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
