

Mutational analysis of the flagellar protein FliG: sites of interaction with FliM and implications for organization of the switch complex
- PMID: 17085573
- PMCID: PMC1797384
- DOI: 10.1128/JB.01281-06
Mutational analysis of the flagellar protein FliG: sites of interaction with FliM and implications for organization of the switch complex
Abstract
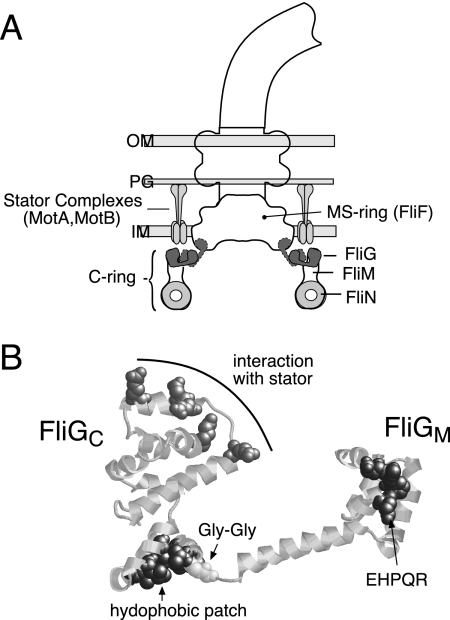
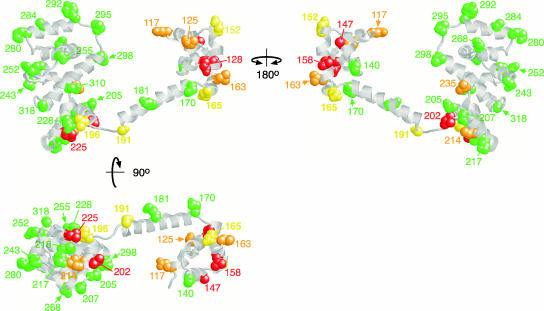
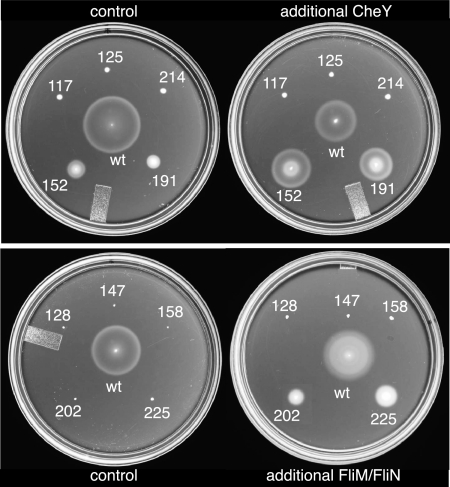
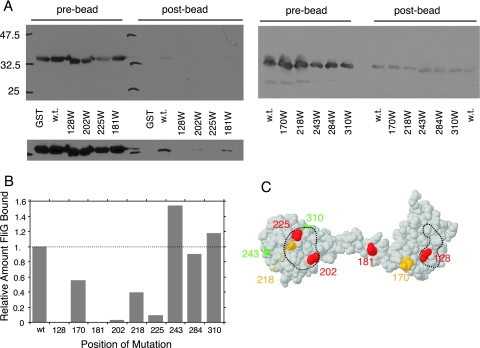
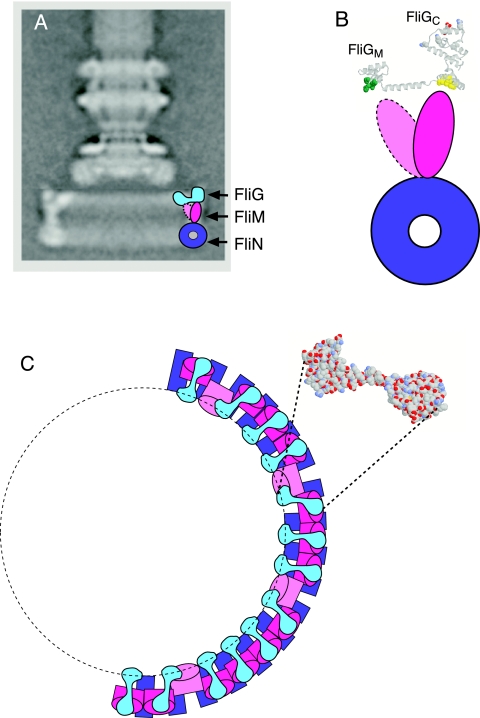
The switch complex at the base of the bacterial flagellum is essential for flagellar assembly, rotation, and switching. In Escherichia coli and Salmonella, the complex contains about 26 copies of FliG, 34 copies of FliM, and more then 100 copies of FliN, together forming the basal body C ring. FliG is involved most directly in motor rotation and is located in the upper (membrane-proximal) part of the C ring. A crystal structure of the middle and C-terminal parts of FliG shows two globular domains connected by an alpha-helix and a short extended segment. The middle domain of FliG has a conserved surface patch formed by the residues EHPQ(125-128) and R(160) (the EHPQR motif), and the C-terminal domain has a conserved surface hydrophobic patch. To examine the functional importance of these and other surface features of FliG, we made mutations in residues distributed over the protein surface and measured the effects on flagellar assembly and function. Mutations preventing flagellar assembly occurred mainly in the vicinity of the EHPQR motif and the hydrophobic patch. Mutations causing aberrant clockwise or counterclockwise motor bias occurred in these same regions and in the waist between the upper and lower parts of the C-terminal domain. Pull-down assays with glutathione S-transferase-FliM showed that FliG interacts with FliM through both the EHPQR motif and the hydrophobic patch. We propose a model for the organization of FliG and FliM subunits that accounts for the FliG-FliM interactions identified here and for the different copy numbers of FliG and FliM in the flagellum.
Figures
Comment in
-
How 34 pegs fit into 26 + 8 holes in the flagellar motor.J Bacteriol. 2007 Jan;189(2):291-3. doi: 10.1128/JB.01556-06. Epub 2006 Nov 3. J Bacteriol. 2007. PMID: 17085552 Free PMC article. No abstract available.
References
-
- Attmannspacher, U., B. Scharf, and R. Schmitt. 2005. Control of speed modulation (chemokinesis) in the unidirectional rotary motor of Sinorhizobium meliloti. Mol. Microbiol. 56:708-718. - PubMed
-
- Berg, H. C. 2003. The rotary motor of bacterial flagella. Annu. Rev. Biochem. 72:19-54. - PubMed
-
- Blair, D. F., and H. C. Berg. 1988. Restoration of torque in defective flagellar motors. Science 242:1678-1681. - PubMed
-
- Block, S. M., and H. C. Berg. 1984. Successive incorporation of force-generating units in the bacterial rotary motor. Nature 309:470-472. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
