

Wza the translocon for E. coli capsular polysaccharides defines a new class of membrane protein
- PMID: 17086202
- PMCID: PMC3315050
- DOI: 10.1038/nature05267
Wza the translocon for E. coli capsular polysaccharides defines a new class of membrane protein
Abstract
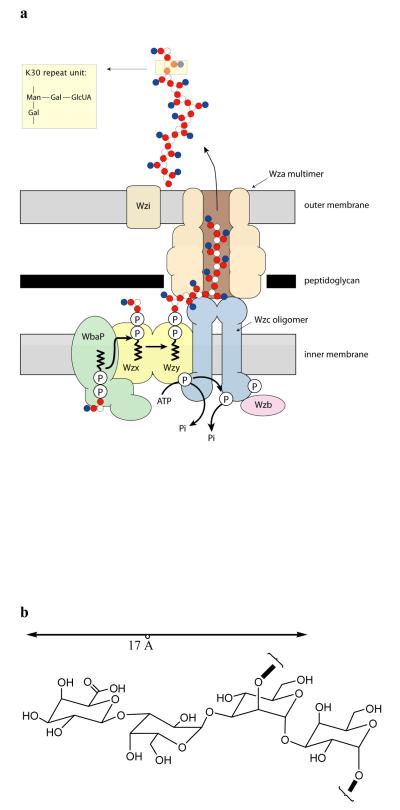
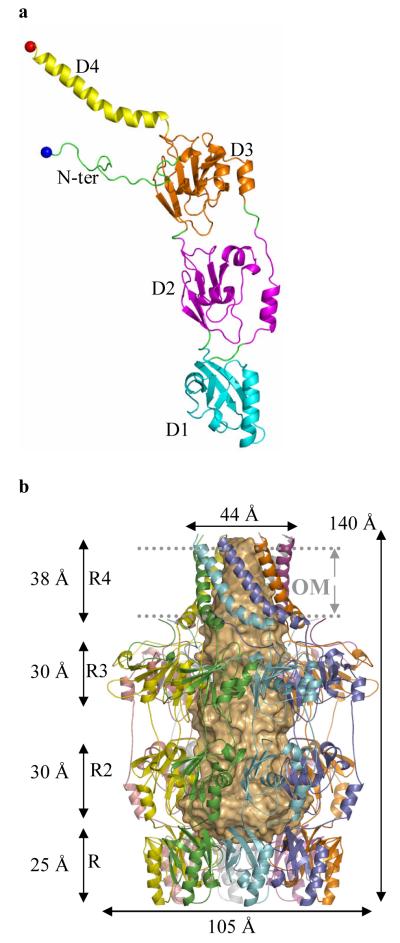
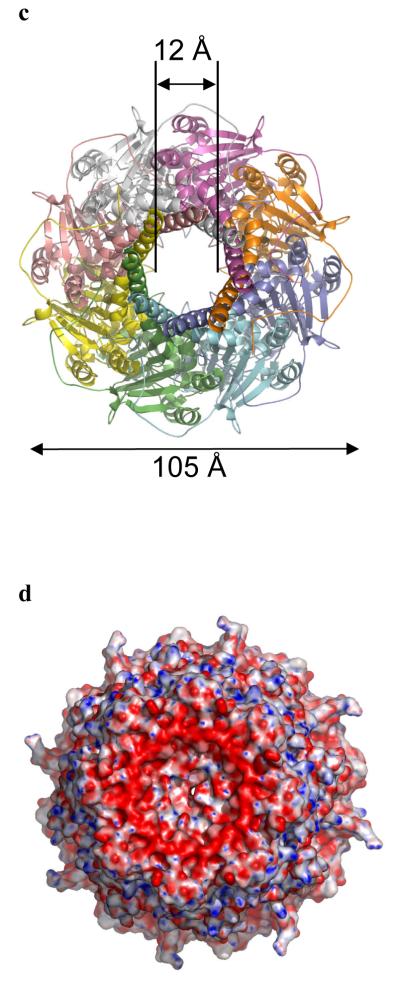
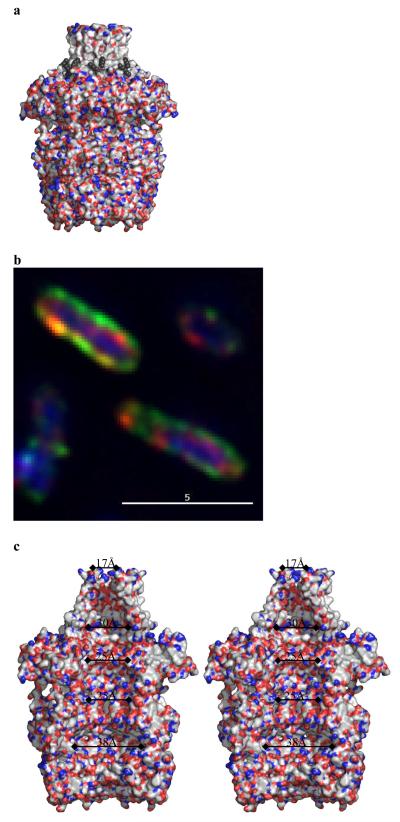
Many types of bacteria produce extracellular polysaccharides (EPSs). Some are secreted polymers and show only limited association with the cell surface, whereas others are firmly attached to the cell surface and form a discrete structural layer, the capsule, which envelopes the cell and allows the bacteria to evade or counteract the host immune system. EPSs have critical roles in bacterial colonization of surfaces, such as epithelia and medical implants; in addition some EPSs have important industrial and biomedical applications in their own right. Here we describe the 2.26 A resolution structure of the 340 kDa octamer of Wza, an integral outer membrane lipoprotein, which is essential for group 1 capsule export in Escherichia coli. The transmembrane region is a novel alpha-helical barrel. The bulk of the Wza structure is located in the periplasm and comprises three novel domains forming a large central cavity. Wza is open to the extracellular environment but closed to the periplasm. We propose a route and mechanism for translocation of the capsular polysaccharide. This work may provide insight into the export of other large polar molecules such as DNA and proteins.
Figures
References
-
- Roberts IS. The biochemistry and genetics of capsular polysaccharide production in bacteria. Annu. Rev. Microbiol. 1996;50:285–315. - PubMed
-
- Hall-Stoodley L, Costerton JW, Stoodley P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004;2:95–108. - PubMed
-
- Fux CA, Costerton JW, Stewart PS, Stoodley P. Survival strategies of infectious biofilms. Trends Microbiol. 2005;13:34–40. - PubMed
-
- Sutherland IW. Novel and established applications of microbial polysaccharides. Trends Biotechnol. 1998;16:41–46. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
