

Rapidly inducible changes in phosphatidylinositol 4,5-bisphosphate levels influence multiple regulatory functions of the lipid in intact living cells
- PMID: 17088424
- PMCID: PMC2064515
- DOI: 10.1083/jcb.200607116
Rapidly inducible changes in phosphatidylinositol 4,5-bisphosphate levels influence multiple regulatory functions of the lipid in intact living cells
Abstract
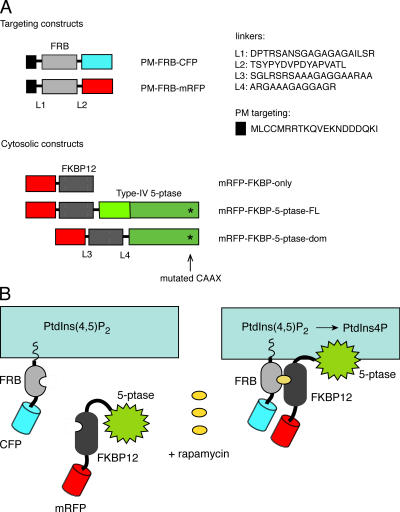
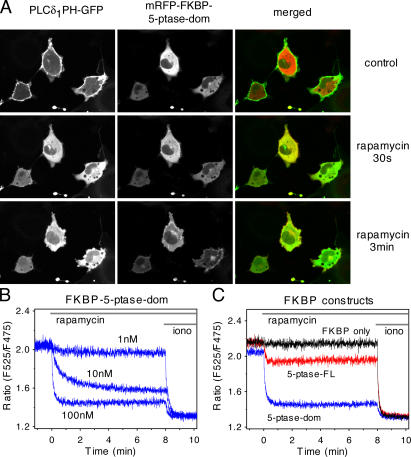
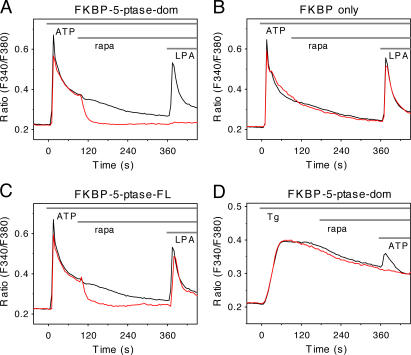
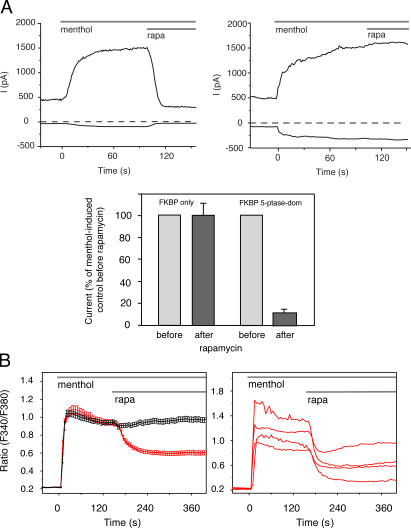
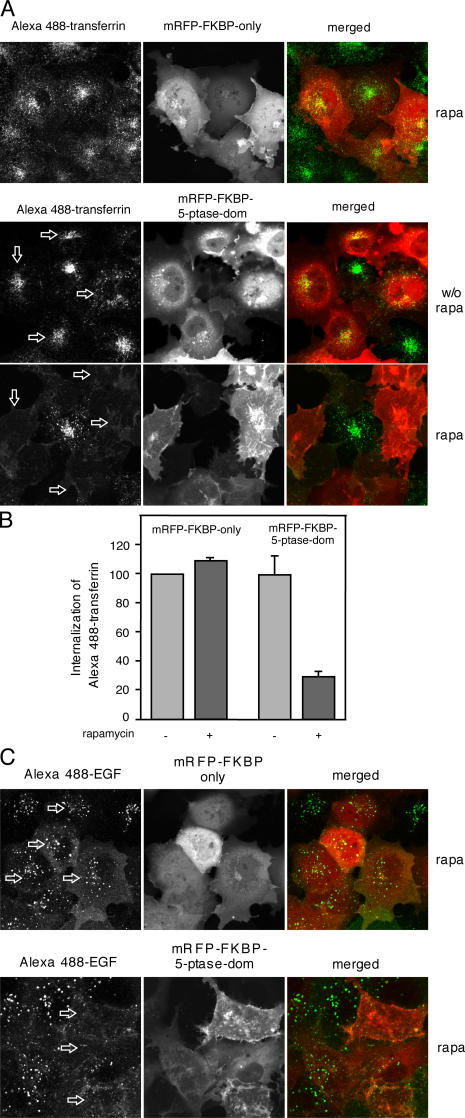
Rapamycin (rapa)-induced heterodimerization of the FRB domain of the mammalian target of rapa and FKBP12 was used to translocate a phosphoinositide 5-phosphatase (5-ptase) enzyme to the plasma membrane (PM) to evoke rapid changes in phosphatidylinositol 4,5-bisphosphate (PtdIns(4,5)P(2)) levels. Rapa-induced PM recruitment of a truncated type IV 5-ptase containing only the 5-ptase domain fused to FKBP12 rapidly decreased PM PtdIns(4,5)P(2) as monitored by the PLCdelta1PH-GFP fusion construct. This decrease was paralleled by rapid termination of the ATP-induced Ca(2+) signal and the prompt inactivation of menthol-activated transient receptor potential melastatin 8 (TRPM8) channels. Depletion of PM PtdIns(4,5)P(2) was associated with a complete blockade of transferrin uptake and inhibition of epidermal growth factor internalization. None of these changes were observed upon rapa-induced translocation of an mRFP-FKBP12 fusion protein that was used as a control. These data demonstrate that rapid inducible depletion of PM PtdIns(4,5)P(2) is a powerful tool to study the multiple regulatory roles of this phospholipid and to study differential sensitivities of various processes to PtdIns(4,5)P(2) depletion.
Figures
References
-
- Berridge, M.J. 1993. Cell signalling: a tale of two messengers. Nature. 365:388–389. - PubMed
-
- Broad, L.M., F.J. Braun, J.P. Lievremont, G.S. Bird, T. Kurosaki, and J.W. Putney Jr. 2001. Role of the phospholipase C-inositol 1,4,5-trisphosphate pathway in calcium release-activated calcium current and capacitative calcium entry. J. Biol. Chem. 276:15945–15952. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
