

Adaptation to ER stress is mediated by differential stabilities of pro-survival and pro-apoptotic mRNAs and proteins
- PMID: 17090218
- PMCID: PMC1634883
- DOI: 10.1371/journal.pbio.0040374
Adaptation to ER stress is mediated by differential stabilities of pro-survival and pro-apoptotic mRNAs and proteins
Abstract
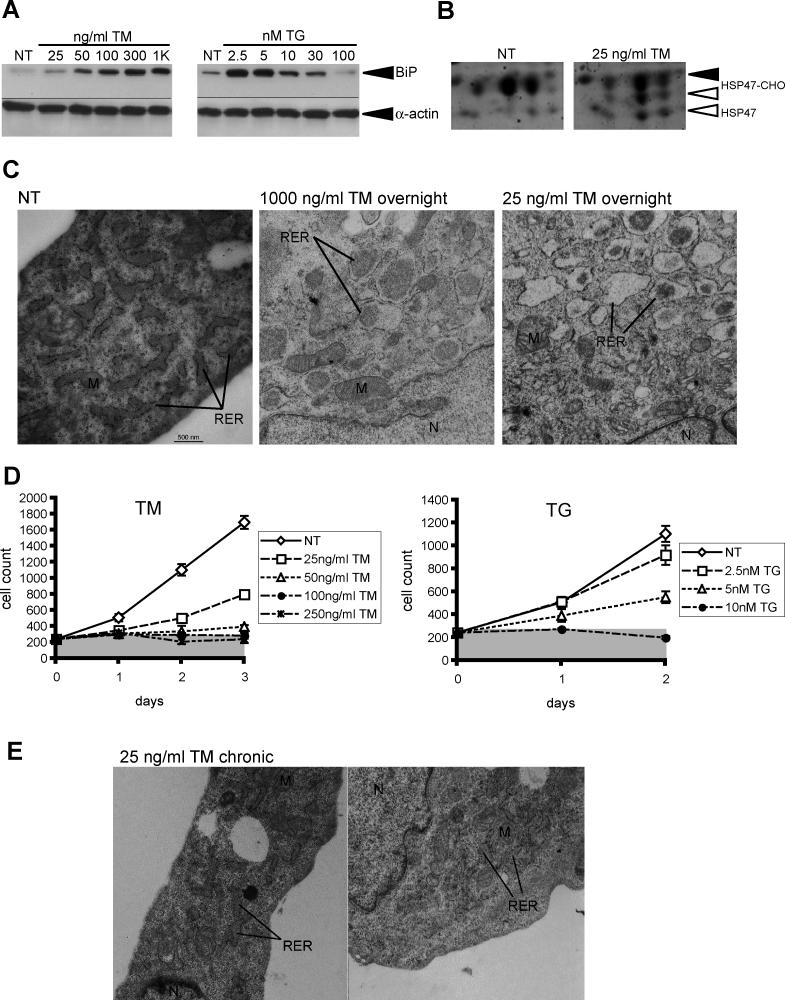
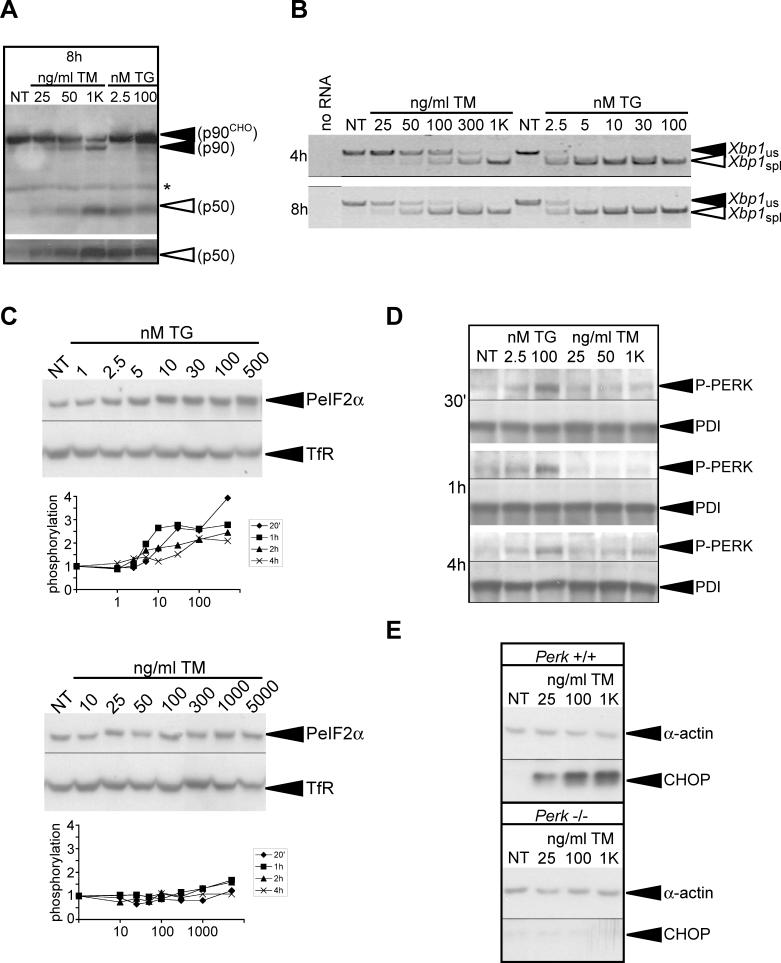
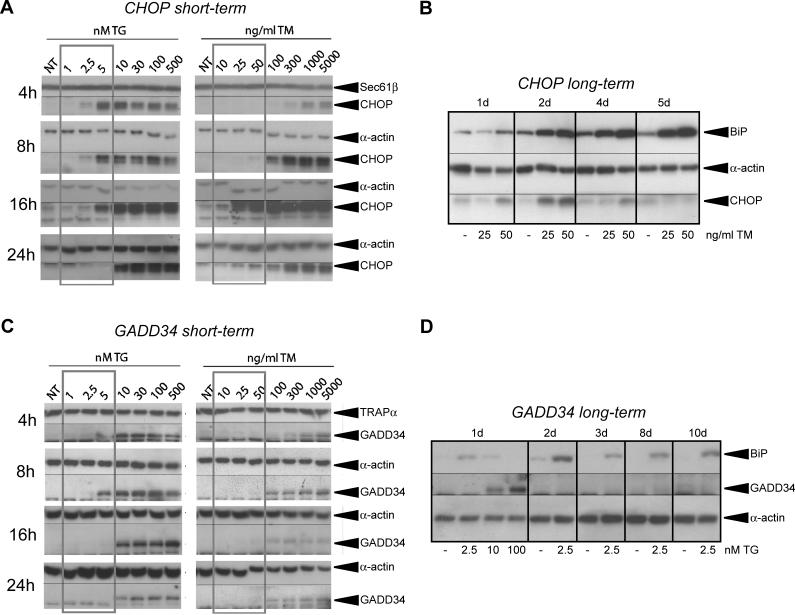
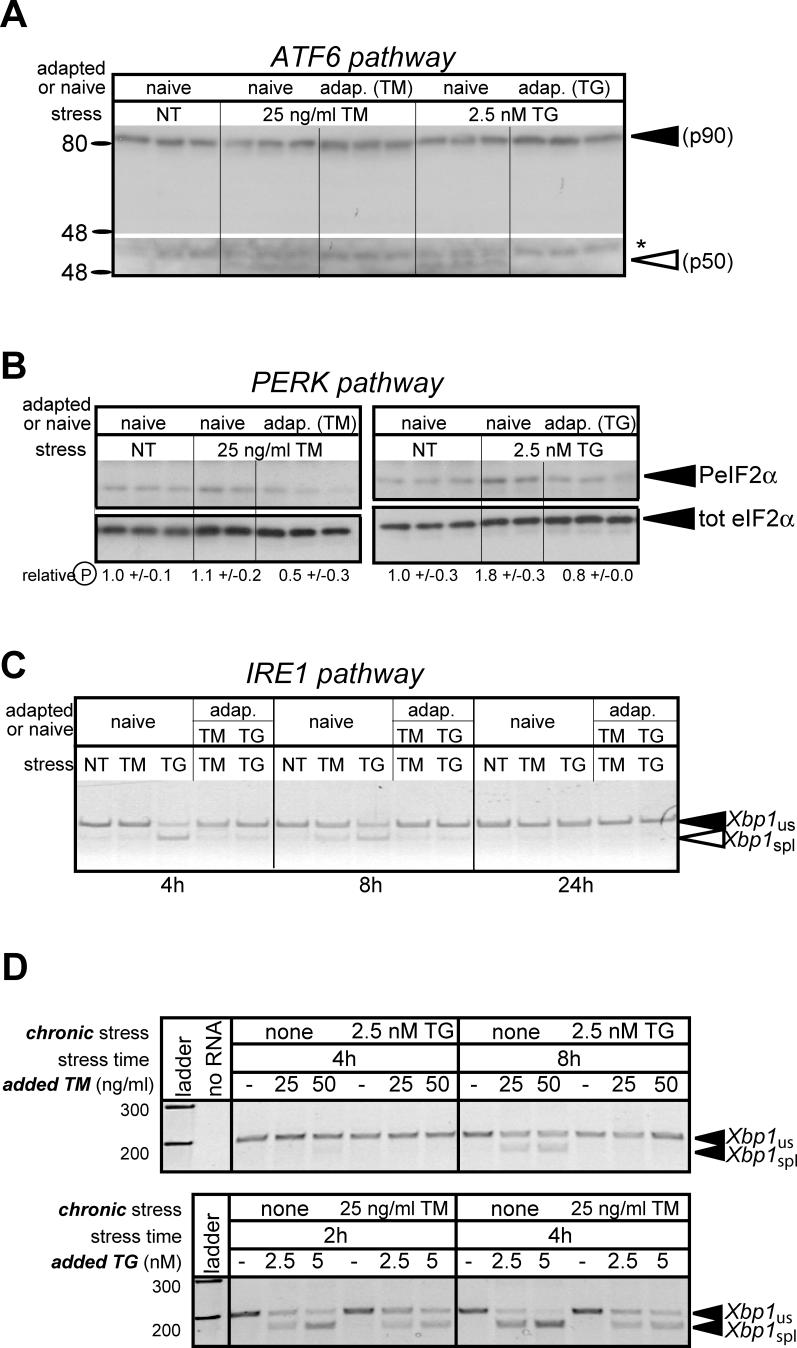
The accumulation of unfolded proteins in the endoplasmic reticulum (ER) activates a signaling cascade known as the unfolded protein response (UPR). Although activation of the UPR is well described, there is little sense of how the response, which initiates both apoptotic and adaptive pathways, can selectively allow for adaptation. Here we describe the reconstitution of an adaptive ER stress response in a cell culture system. Monitoring the activation and maintenance of representative UPR gene expression pathways that facilitate either adaptation or apoptosis, we demonstrate that mild ER stress activates all UPR sensors. However, survival is favored during mild stress as a consequence of the intrinsic instabilities of mRNAs and proteins that promote apoptosis compared to those that facilitate protein folding and adaptation. As a consequence, the expression of apoptotic proteins is short-lived as cells adapt to stress. We provide evidence that the selective persistence of ER chaperone expression is also applicable to at least one instance of genetic ER stress. This work provides new insight into how a stress response pathway can be structured to allow cells to avert death as they adapt. It underscores the contribution of posttranscriptional and posttranslational mechanisms in influencing this outcome.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures
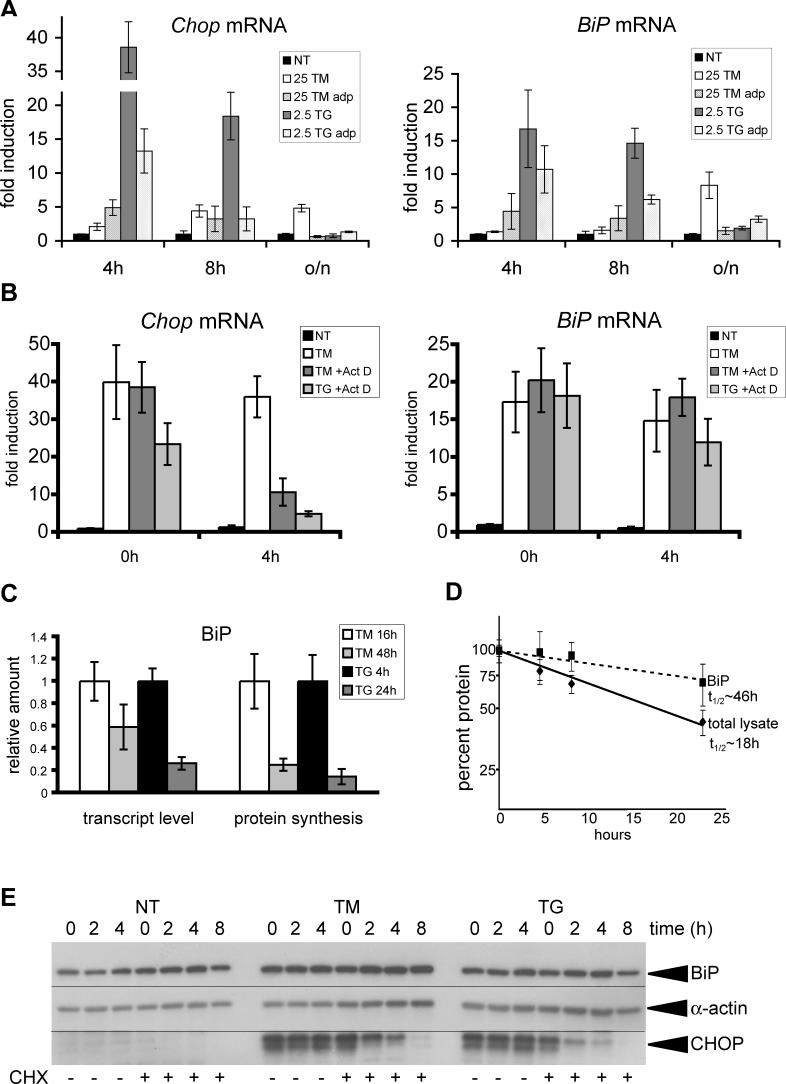
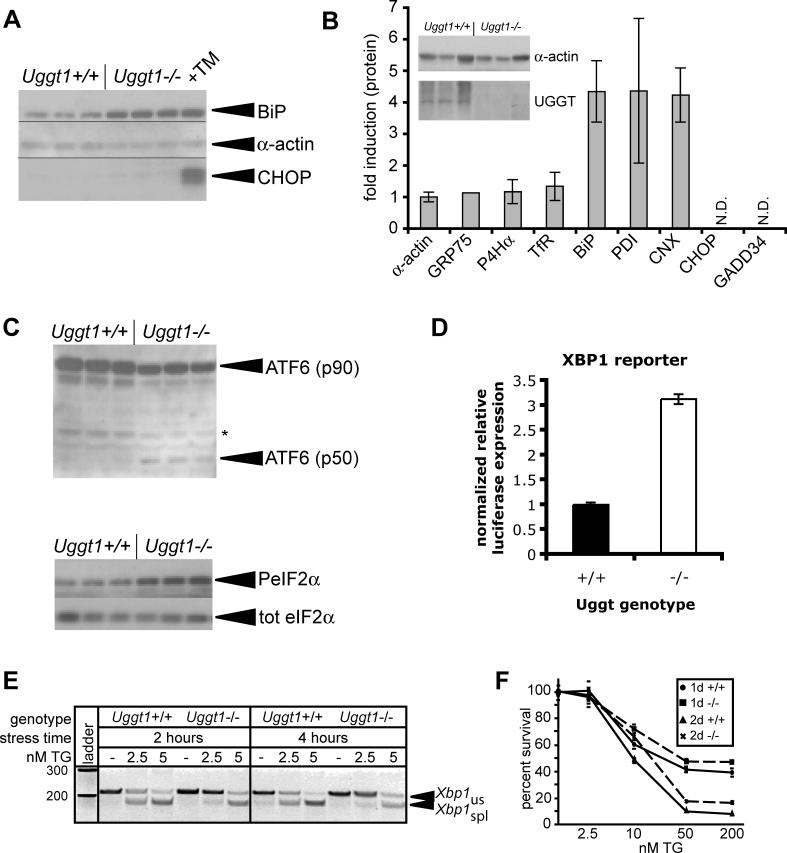
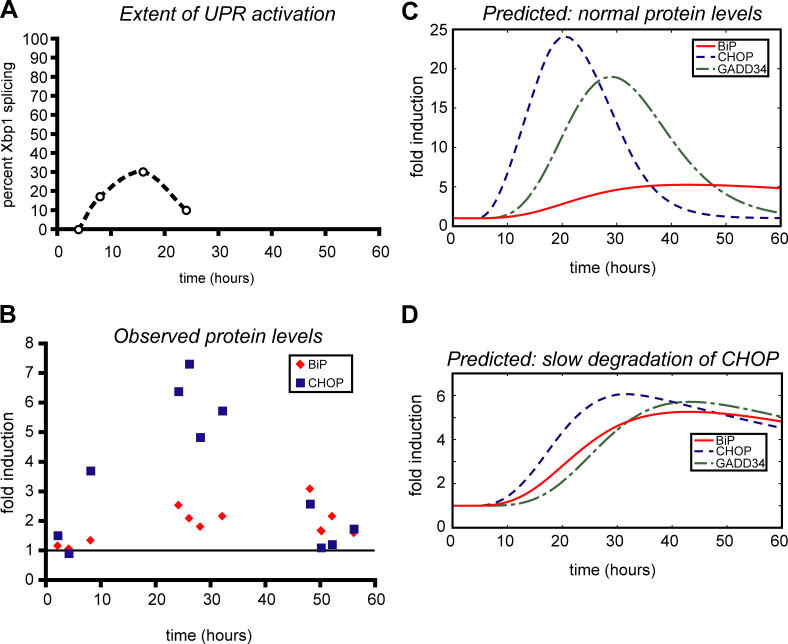
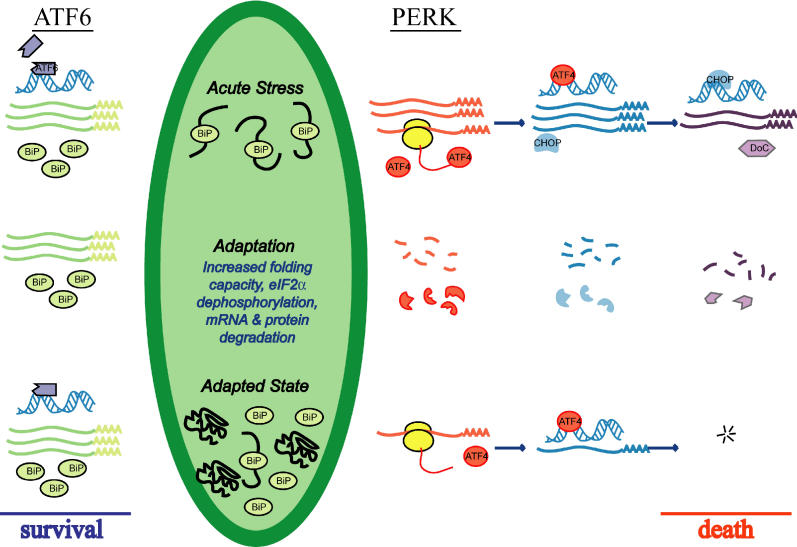
Comment in
-
Apoptosis versus survival: when in doubt, choose both.PLoS Biol. 2006 Nov;4(11):e408. doi: 10.1371/journal.pbio.0040408. Epub 2006 Nov 7. PLoS Biol. 2006. PMID: 20076503 Free PMC article. No abstract available.
References
-
- Rutkowski DT, Kaufman RJ. A trip to the ER: Coping with stress. Trends Cell Biol. 2004;14:20–28. - PubMed
-
- Bertolotti A, Zhang Y, Hendershot LM, Harding HP, Ron D. Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat Cell Biol. 2000;2:326–332. - PubMed
-
- Liu CY, Schroder M, Kaufman RJ. Ligand-independent dimerization activates the stress response kinases IRE1 and PERK in the lumen of the endoplasmic reticulum. J Biol Chem. 2000;275:24881–24885. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
